 |
Главная |


Явление сцепленного наследования
|
из
5.00
|
Явление сцепленного наследования изучено Т. Морганом, который установил, что материальной основой сцепления является хромосома (хромосомная теория наследственности). Суть сцепленного наследования как нарушение сцепления, происходящего в результате перекреста хромосом, или кроссинговера, необходимо обратить внимание на биологический смысл этого феномена. При перекресте хромосом происходит обмен идентичными участками между гомологичными хромосомами, а значит, возникают новые комбинации генов (как аллельных, так и неаллельных). Обсуждая вопрос о природе изменчивости живых организмов.
Группы сцепления.
Число генов у каждого организма гораздо больше числа хромосом. Следовательно, в одной хромосоме расположено много генов. Ученые установили, что гены, расположенные в одной хромосоме, наследуются совместно, или сцеплено. Группы генов, расположенные в одной хромосоме, называют группами сцепления. Сцепленные гены расположены в хромосоме в линейном порядке. Число групп сцепления у генетически хорошо изученных объектов равно числу пар хромосом, т.е. гаплоидному числу хромосом. У человека 23 пары хромосом и 23 группы сцепления.
Кроссинговер.
Если гены находятся в одной хромосоме и всегда передаются вместе, говорят о полном сцеплении. Чаще встречается неполное сцепление. Нарушения сцепления объясняется кроссинговером, который является обменом идентичных участков гомологичных хромосом, в которых расположены аллельные гены. Запись 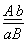 означает, что в одной аутосоме находится доминантный ген 1-ой пары альтернативных признаков и рецессивный ген 2-ой. А в другой аутосоме наоборот. В половых хромосомах
означает, что в одной аутосоме находится доминантный ген 1-ой пары альтернативных признаков и рецессивный ген 2-ой. А в другой аутосоме наоборот. В половых хромосомах 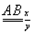 y-хромосома не несет этих генов. Кроме сцепления генов, здесь идет сцепление с полом.
y-хромосома не несет этих генов. Кроме сцепления генов, здесь идет сцепление с полом.
Кроссовер - гамета, которая претерпела процесс кроссинговера. Частота вступления генов в кроссинговер прямо пропорциональна расстоянию между ними, поэтому число гамет с новыми комбинированными формами будет зависеть от расстояния между генами. Расстояние вычисляется в морганидах, но если речь идет о кроссинговере, то расстояние вычисляется в процентах
 .
.
Одной морганиде соответствует 1% образования гамет, в которых гомологичные хромосомы обмениваются своими участками. 50М - максимальное расстояние между генами, на котором возможен кроссинговер. Если гены расположены друг от друга на расстоянии, большем 50М, то наблюдается явление независимого наследования. На основании частот кроссинговера строится карта группы сцепления.
Кроссинговер может происходить не только во время мейоза, но и митоза, тогда его называют митотическим кроссинговером. Частота митотического кроссинговера значительно ниже мейотического. Тем не менее, его также можно использовать для генетического картирования.
Мейотический кроссинговер осуществляется после того, как гомологичные хромосомы в зиготной стадии профазы I соединяются в пары, образуя биваленты. В профазе I каждая хромосома представлена двумя сестринскими хроматидами, и перекрест происходит между хроматидами.
Приняв положения, что 1) генов в хромосоме может быть много, 2) гены расположены в хромосоме в линейном порядке, 3) каждая аллельная пара занимает определенные и идентичные локусы в гомологичных хромосомах, Т. Морган допустил, что перекрест между хроматидами гомологичных хромосом может происходить одновременно в нескольких точках кроссинговер, происходящий лишь в одном месте, называют одиночным кроссинговером, в двух точках одновременно - двойным, в трех - тройным и т.д., т.е. кроссинговер может быть множественным.
Пусть, например, в гомологичной паре хромосом содержатся три пары аллелей в гетерозиготном состоянии  . Тогда перекрест, произошедший только в участке между генами А и В или между В и С, будет одинарным. В результате одинарного перекреста возникают в каждом случае только две кроссоверные хромосомы aBC и Abc или Abc и aBC.
. Тогда перекрест, произошедший только в участке между генами А и В или между В и С, будет одинарным. В результате одинарного перекреста возникают в каждом случае только две кроссоверные хромосомы aBC и Abc или Abc и aBC.
Каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках. Таким образом, двойные кроссинговеры сокращают регистрируемое расстояние между генами.
Вместе с тем между обменами на соседних участках хромосом существует взаимовлияние, названное интерференцией. Такое взаимовлияние можно выразить количественно. Для этого составляют реально наблюдаемую частоту двойных кроссинговеров с частотой, теоретически ожидаемой на основе предположения о том, что обмены на соседних участках происходят независимо друг от друга. Степень и характер интерференции измеряется величиной коинциденции (С). Коинциденцию оценивают как частное от деления реально наблюдаемой частоты двойных кроссоверов на теоретически ожидаемую частоту двойных кроссоверов. Последнюю величину получают, перемножая частоты кроссинговера на соседних участках.
Величину интерференции (I) определяют по формуле I=1-C. Если С<1, то интерференция положительная, т.е. одинаковый обмен препятствует обмену на соседнем участке хромосомы. Если С>1, то интерференция отрицательная, т.е. один обмен как бы стимулирует дополнительные обмены на соседних участках.
В действительности существует только положительная интерференция при реципрокной рекомбинации - кроссинговере, а кажущееся неслучайным совпадение двух и более обменов, характерное для очень коротких расстояний - результат нереципрокных событий при рекомбинации.
Таким образом, при карплеровании генов в группах сцепления на основе изучения частот рекомбинации необходимо учитывать две противоположные тенденции. Двойные обмены “сокращают” расстояния между генами, и интерференция препятствует множественным обменам, вероятность которых увеличивается с расстоянием.
В обобщенном виде зависимость частоты рекомбинации от реального расстояния с учетом множественных обменов описывает функция Дж. Холдэйна

где rf - картирующая функция (в нашем случае - это частота учитываемых кроссинговеров), d - реальное расстояние, на котором происходят обмены, e - основание натурального логарифма.
При изучении множественных обменов и интерференции между ними используют тетрадный анализ. Для этого рассматривают тригибридное скрещивание (ABCxabc) по сцепленным генам. Учитывая, что кроссинговер происходит на стадии 4-х хроматид, возможны три типа двойных обменов. Это двойные двухроматидные обмены, двойные треххроматидные обмены и двойные четыреххроматидные обмены только между несестринскими хроматидами, последствия которых генетически различимы.
|
из
5.00
|
Обсуждение в статье: Явление сцепленного наследования |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы