 |
Главная |


Законы раздражения возбудимых тканей
|
из
5.00
|
Лекция 1
ОБЩИЕ ЗАКОНОМЕРНОСТИ РЕАГИРОВАНИЯ ЖИВОЙ МАТЕРИИ
План:
1. Биоэлектрические явления в возбудимых тканях. 1
2. Мембранный потенциал. 3
3. Потенциал действия. 6
4. Законы раздражения возбудимых тканей. 9
Биоэлектрические явления в возбудимых тканях
Способность адаптироваться к постоянно изменяющимся условиям внешней среды является одним из основных признаков живых систем. В основе приспособительных реакций организма лежит раздражимость – способность реагировать на действие различных факторов изменением структуры и функций. Раздражимостью обладают все ткани животных и растительных организмов. В процессе эволюции происходила постепенная дифференциация тканей, участвующих в приспособительной деятельности организма. Раздражимость этих тканей достигла наивысшего развития и трансформировалась в новое свойство – возбудимость. Под этим термином понимают способность ряда тканей (нервной, мышечной, железистой) отвечать на раздражение генерацией процесса возбуждения. Возбуждение – это сложный физиологический процесс временной деполяризации мембраны клеток, который проявляется специализированной реакцией ткани (проведение нервного импульса, сокращение мышцы, отделение секрета железой и т. д.). Возбудимостью обладают нервная, мышечная и секреторная ткани, которые называют возбудимыми тканями. Возбудимость различных тканей неодинакова. Ее величину оценивают по порогу раздражения – минимальной силе раздражителя, которая способна вызвать возбуждение. Менее сильные раздражители называются подпороговыми, а более сильные – сверхпороговыми.
Раздражителями, вызывающими возбуждение, могут быть любые внешние (действующие из окружающей среды) или внутренние (возникающие в самом организме) воздействия. Все раздражители по их природе можно разделить на три группы: физические (механические, электрические, температурные, звуковые, световые), химические (щелочи, кислоты и другие химические вещества, в том числе и лекарственные) и биологические (вирусы, бактерии, насекомые и другие живые существа).
По степени приспособленности биологических структур к их восприятию раздражители можно разделить на адекватные и неадекватные. Адекватными называются раздражители, к восприятию которых биологическая структура специально приспособлена в процессе эволюции. Например, адекватным раздражителем для фоторецепторов является свет, для барорецепторов – изменение давления, для мышц – нервный импульс. Неадекватными называются такие раздражители, которые действуют на структуру, специально не приспособленную для их восприятия. Например, мышца может сокращаться под влиянием механического, теплового, электрического раздражений, хотя адекватным раздражителем для нее является нервный импульс. Пороговая сила неадекватных раздражителей во много раз превышает пороговую силу адекватных.
Возбуждение представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и кратковременное изменение электрического потенциала мембраны.
Первые исследования электрической активности живых тканей были проведены Л. Гальвани. Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенной на медном крючке, при соприкосновении с железными перилами балкона (первый опыт Гальвани). На основании этих наблюдений им был сделал вывод, что сокращение лапок вызвано «животным электричеством», которое возникает в спинном мозге и передается по металлическим проводникам (крючку и перилам) к мышцам.
Физик А. Вольта, повторив этот опыт, пришел к другому заключению. Источником тока, по его мнению, является не спинной мозг и «животное электричество», а разность потенциалов, образующаяся в месте контакта разнородных металлов – меди и железа, а нервно-мышечный препарат лягушки является лишь проводником электричества. В ответ на эти возражения Л. Гальвани усовершенствовал опыт, исключив из него металлы. Он препарировал седалищный нерв вдоль бедра лапки лягушки, затем набрасывал нерв на мышцы голени, что вызывало сокращение мышцы (второй опыт Гальвани), тем самым доказав существование «животного электричества».
Позднее Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок – положительный. При набрасывании нерва между поврежденным и неповрежденным участками мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя, или током повреждения. Так было показано, что наружная поверхность мышечных клеток заряжена положительно по отношению к внутреннему содержимому.
Мембранный потенциал
В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая называется мембранным потенциалом (МП), или, если это клетка возбудимой ткани, – потенциалом покоя. Так как внутренняя сторона мембраны заряжена отрицательно по отношению к наружной, то, принимая потенциал наружного раствора за нуль, МП записывают со знаком «минус». Его величина у разных клеток колеблется от минус 30 до минус 100 мВ.
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю.Бернштейном (1902). Исходя из того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он показал, что величину мембранного потенциала можно определить, используя формулу Нернста:
 ,
,
где Ем – разность потенциалов между внутренней и наружной сторонами мембраны; Ек– равновесный потенциал для ионов калия; R – газовая постоянная; Т – абсолютная температура; n – валентность иона; F – число Фарадея; [К+]вн – внутренняя и [К+]н – наружная концентрация ионов калия.
В 1949-1952 гг. А.Ходжкин, Э.Хаксли, Б.Катц создали современную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциалозависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией по обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле:

где РК, PNa, РСl – проницаемость для ионов калия, натрия и хлора.
Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т.е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То, что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.
Если раздражать нейрон через электрод, находящийся в цитоплазме, кратковременными импульсами деполяризующего электрического тока различной величины, то, регистрируя через другой электрод изменения мембранного потенциала, можно наблюдать следующие биоэлектрические реакции: электротонический потенциал, локальный ответ и потенциал действия (рис. 1).

Рис. 1. Изменение мембранного потенциала под влиянием деполяризующих и гиперполяризующих раздражений: a – электротонический потенциал; б – локальный ответ; в – потенциал действия; г – гиперполяризация; д – раздражения.
Если наносятся раздражения, величина которых не превышает 0,5 величины порогового раздражения, то деполяризация мембраны наблюдается только во время действия раздражителя. Это пассивная электротоническая деполяризация (электротонический потенциал). Развитие и исчезновение электротонического потенциала происходит по экспоненте (возрастает) и определяется параметрами раздражающего тока, а также свойствами мембраны (ее сопротивлением и емкостью). Во время развития электротонического потенциала проницаемость мембраны для ионов практически не изменяется.
Локальный ответ. При увеличении амплитуды подпороговых раздражений от 0,5 до 0,9 пороговой величины развитие деполяризации мембраны происходит не прямолинейно, а по S-образной кривой. Деполяризация продолжает нарастать и после прекращения раздражения, а затем сравнительно медленно исчезает. Этот процесс получил название локального ответа. Локальный ответ имеет следующие свойства:
1) возникает при действии подпороговых раздражителей;
2) находится в градуальной зависимости от силы стимула (не подчиняется закону «все или ничего»); локализуется в месте действия раздражителя и не способен к распространению на большие расстояния;
3) может распространяться лишь локально, при этом его амплитуда быстро уменьшается;
4) локальные ответы способны суммироваться, что приводит к увеличению деполяризации мембраны.
В период развития локального ответа возрастает поток ионов натрия в клетку, что повышает ее возбудимость. Локальный ответ является экспериментальным феноменом, однако по перечисленным выше свойствам он близок к таким явлениям, как процесс местного нераспространяющегося возбуждения и возбуждающего постсинаптического потенциала (ВПСП), который возникает под влиянием деполяризующего действия возбуждающих медиаторов.
Потенциал действия
Потенциал действия (ПД) возникает на мембранах возбудимых клеток под влиянием раздражителя пороговой или сверхпороговой величины, который увеличивает проницаемость мембраны для ионов натрия. Ионы натрия начинают входить внутрь клетки, что приводит к уменьшению величины мембранного потенциала – деполяризации мембраны. При уменьшении МП до критического уровня деполяризации открываются потенциалозависимые каналы для натрия и проницаемость мембраны для этих ионов увеличивается в 500 раз (превышая проницаемость для ионов калия в 20 раз). В результате проникновения ионов натрия в цитоплазму и их взаимодействия с анионами разность потенциалов на мембране исчезает, а затем происходит перезарядка клеточной мембраны (инверсия заряда, овершут) – внутренняя поверхность мембраны заряжается положительно по отношению к наружной (на 30-50 мВ), после чего закрываются натриевые каналы и открываются потенциалозависимые калиевые каналы. В результате выхода калия из клетки начинается процесс восстановления исходного уровня мембранного потенциала покоя – реполяризация мембраны. Если такое повышение проводимости для калия предотвратить введением тетраэтиламмония, который избирательно блокирует калиевые каналы, мембрана реполяризуется гораздо медленнее. Натриевые каналы можно блокировать тетродотоксином и разблокировать последующим введением фермента проназы, который расщепляет белки.
Таким образом, в основе возбуждения (генерации ПД) лежит повышение проводимости мембраны для натрия, вызываемое ее деполяризацией до порогового (критического) уровня.
В потенциале действия различают следующие фазы:
1. Предспайк – процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
2. Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
3. Отрицательный следовой потенциал – от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
4. Положительный следовой потенциал – увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
При развитии потенциала действия происходят фазные изменения возбудимости ткани (рис. 2). Состоянию исходной поляризации мембраны (мембранный потенциал покоя) соответствует нормальный уровень возбудимости. В период предспайка возбудимость ткани повышена. Эта фаза возбудимости получила название повышенной возбудимости (первичной экзальтации). В это время мембранный потенциал приближается к критическому уровню деполяризации, поэтому дополнительный стимул, даже если он меньше порогового, может довести мембрану до критического уровня деполяризации. В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны и она утрачивает способность отвечать возбуждением на раздражители даже сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности (абсолютной невозбудимости). Она длится до конца перезарядки мембраны и возникает в связи с тем, что натриевые каналы инактивируются.

Рис.2. Соотношение одиночного цикла возбуждения (А) и фаз возбудимости (Б).
Для А: а – мембранный потенциал покоя; б – локальный ответ или ВПСП; в – восходящая фаза потенциала действия (деполяризация и инверсии); г – нисходящая фаза потенциала действия (реполяризация); д – отрицательный следовой потенциал (следовая деполяризация); е – положительный следовой потенциал (следовая гиперполяризация).
Для Б: а – исходный уровень возбудимости; б – фаза повышенной возбудимости; в – фаза абсолютной рефрактерности; г – фаза относительной рефрактерности; д – фаза супернормальной возбудимости; е – фаза субнормальной возбудимости.
После окончания фазы перезарядки мембраны возбудимость ее постепенно восстанавливается до исходного уровня – фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны, достигая величины критического уровня деполяризации. Так как в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя.
Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых. Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости (фаза вторичной экзальтации). Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации по сравнению с состоянием покоя (исходной поляризацией), то порог раздражения снижен и новое возбуждение может возникнуть при действии раздражителей подпороговой силы.
В период развития положительного следового потенциала возбудимость ткани понижена – фаза субнормальной возбудимости (вторичной рефрактерности). В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации мембраны), удаляясь от критического уровня деполяризации, порог раздражения повышается и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Рефрактерность мембраны является следствием того, что натриевый канал состоит из собственно канала (транспортной части) и воротного механизма, который управляется электрическим полем мембраны. В канале предполагают наличие двух типов «ворот» – быстрых активационных (m) и медленных инактивационных (h). «Ворота» могут быть полностью открыты или закрыты, например, в натриевом канале в состоянии покоя «ворота» m закрыты, а «ворота» h – открыты. При уменьшении заряда мембраны (деполяризации) в начальный момент «ворота» m и h открыты – канал способен проводить ионы. Через открытые каналы ионы движутся по концентрационному и электрохимическому градиенту. Затем инактивационные «ворота» закрываются, т.е. канал инактивируется. По мере восстановления МП инактивационные «ворота» медленно открываются, а активационные быстро закрываются и канал возвращается в исходное состояние. Следовая гиперполяризация мембраны может возникать вследствие трех причин: во-первых, продолжающимся выходом ионов калия; во-вторых, открытием каналов для хлора и поступлением этих ионов в клетку; в-третьих, усиленной работой натрий-калиевого насоса.
Законы раздражения возбудимых тканей
Эти законы отражают определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани. К законам раздражения относятся: закон силы, закон раздражения Дюбуа-Реймона (аккомодации), закон силы-времени (силы-длительности).
Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции. В соответствии с этим законом функционирует скелетная мышца. Амплитуда ее сокращений постепенно увеличивается с увеличением силы раздражителя вплоть до достижения максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. На пороговые раздражители отвечают только волокна, имеющие самую высокую возбудимость, амплитуда мышечного сокращения при этом минимальна. Увеличение силы раздражителя приводит к постепенному вовлечению волокон, имеющих меньшую возбудимость, поэтому амплитуда сокращения мышцы усиливается. Когда в реакции участвуют все мышечные волокна данной мышцы, дальнейшее повышение силы раздражителя не приводит к увеличению амплитуды сокращения.
Закон раздражения Дюбуа-Реймона (аккомодации): стимулирующее действие постоянного тока зависит не только от абсолютной величины силы тока, но и от скорости нарастания тока во времени. При действии медленно нарастающего тока возбуждение не возникает, так как происходит приспособление возбудимой ткани к действию этого раздражителя, что получило название аккомодации. Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения ПД не возникает, так как деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого – к окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога генерации ПД. Способность к аккомодации различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
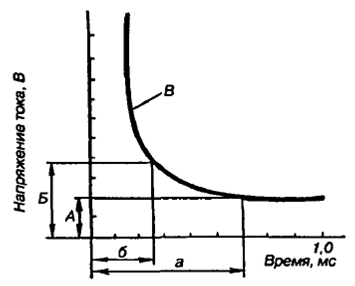
Рис.3. Зависимость между силой тока и временем его действия: А – реобаза; Б – удвоенная реобаза; В – кривая силы времени; а – полезное время действия тока; б – хронаксия
Закон силы-времени: раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать на возбудимые ткани, чтобы вызвать возбуждение (рис.3). Исследования зависимости силы-длительности показали, что она имеет гиперболический характер. Ток меньше некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Причиной такой зависимости является мембранная емкость. Очень «короткие» токи не успевают разрядить эту емкость до критического уровня деполяризации. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого ток, равный реобазе, вызывает возбуждение, называется полезным временем. Хронаксия – минимальное время, в течение которого ток, равный двум реобазам, вызывает ответную реакцию.
Литература
1. Физиология человека / Под ред. Покровского В.М., Коротько Г.Ф. – М.: Медицина, 2003. – 656 с.
2. Филимонов В.И. Руководство по общей и клинической физиологии. – М.: Медицинское информационное агентство, 2002. – 958 с.
3. Фундаментальная и клиническая физиология / Под ред. А.Г.Камкина, А.А.Каменского. – М.: Academia, 2004. – 1072 с.
|
из
5.00
|
Обсуждение в статье: Законы раздражения возбудимых тканей |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы