 |
Главная |


Полиморфизм двойной спирали ДНК
|
из
5.00
|


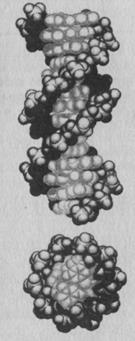

 Модель ДНК Уотсона и Крика описывает структуру одной из нескольких форм двойной спирали. Эта основная форма ДНК, в которой большая часть ее молекул существует в клетке, была названа В-формой.
Модель ДНК Уотсона и Крика описывает структуру одной из нескольких форм двойной спирали. Эта основная форма ДНК, в которой большая часть ее молекул существует в клетке, была названа В-формой.
|
|
|
и сверху): объемные модели
|
Показана зигзагообразная линия, соединяющая фосфатные группы в Z-ДНК.
После остатков гуанина зигзагообразная линия идет вертикально, после остатков цитозина – горизонтально
При понижении влажности В-форма ДНК переходит в А-форму
(рис. 14). Изменение конформации углеводного остатка приводит к уменьшению расстояния между нуклеотидными парами вдоль оси спирали (примерно до 25 нм при 11 нуклеотидных остатках на виток спирали). Диаметр спирали увеличивается, изменяется ширина бороздок, пары оснований смещаются к периферии спирали. Вследствие этого внутри спирали возникает полость диаметром около 0,40 нм. А- и В-формы ДНК – это правозакрученные спирали.
При высокой концентрации солей или добавлении спирта спираль В-типа переходит в левозакрученную Z-форму (рис. 15). Левая спираль была обнаружена у полинуклеотида с чередующейся последовательностью нуклеотидов гуанин-цитозин. Повторяющаяся единица в Z-форме состоит из двух пар нуклеотидов (Г-Ц), в результате линия, соединяющая фосфатные группы, через каждые две пары имеет излом и принимает зигзагообразный вид (отсюда название Z-формы).
Функции ДНК
1. ДНК является носителем генетической информации. Функция обеспечивается фактом существования генетического кода.
2. Воспроизведение и передача генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
3. Реализация генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов. Функция обеспечивается процессами транскрипции и трансляции.
Структура РНК
Рибонуклеиновые кислоты (РНК) представляют собой линейные полинуклеотиды с общими для нуклеиновых кислот принципами организации:
· состоят из четырех видов нуклеотидов, каждый из которых включает азотистое основание (аденин, урацил, гуанин, цитозин), пентозу (рибозу) и фосфатный остаток;
· нуклеотиды связаны в цепь с помощью 3′-5′-фосфодиэфирных связей;
· полинуклеотидные цепи полярны, т. е. имеют различимые 5′- и 3′-концы.
Таблица 1
Отличия РНК и ДНК
| Нуклеиновая кислота Признак | РНК | ДНК |
| Сахар | Рибоза | Дезоксирибоза |
| Азотистые основания | А, У, Г, Ц | А, Т, Г, Ц |
| Количество цепей в молекуле | 99,99% одноцепочечная, 0,01% двухцепочечная | 99,99% двойная спираль, 0,01% одноцепочечная |
| Форма молекулы | Линейные молекулы | Большинство двухцепочечных – линейные, часть – кольцевые |
Главное отличие РНК от ДНК состоит в том, что молекулы РНК являются не двух-, а одноцепочечными. Причиной этому служат следующие особенности первичной структуры РНК:
1. Пентоза в РНК – не дезоксирибоза, а рибоза, которая содержит дополнительную гидроксигруппу. Последняя делает двухцепочечную структуру менее устойчивой.
2. Среди четырех главных (мажорных) азотистых оснований вместо тимина содержится урацил, отличающийся от тимина лишь отсутствием метильной группы у 5-го атома углерода (рис. 12). Благодаря этому уменьшается сила гидрофобного взаимодействия в комплементарной паре А-У. Это снижает вероятность образования устойчивых двухцепочечных молекул.
3. В РНК высоко содержание так называемых минорных оснований. Среди них – дигидроуридин (в урациле нет одной двойной связи), псевдоуридин (урацил иначе, чем обычно, связан с рибозой), диметиладенин и диметилгуанин (в азотистых основаниях – по две дополнительных метильных группы) и многие другие. Почти все эти основания не могут участвовать в комплементарных взаимодействиях.
Перечисленные отличия строения РНК от ДНК имеют большое биологическое значение, так как свою функцию молекулы РНК способны выполнять только в одноцепочечном состоянии.
Общие черты вторичной структуры РНК
Молекулы большинства природных РНК построены из одной полинуклеотидной цепи. Вместе с тем в некоторых участках цепь РНК может образовывать петли, или «шпильки», с двухцепочечной структурой
(рис. 16А). Эта структура стабилизирована взаимодействием оснований в парах А-У, Г-Ц. Однако могут образовываться и «неправильные» пары (например, Г-У), а в некоторых участках «шпильки» вообще не происходит никакого взаимодействия. В составе таких петель может содержаться до 50% всех нуклеотидов.
Третичная структура РНК
|
 В физиологических условиях однотяжевые РНК характеризуются компактной и упорядоченной третичной структурой, которая возникает за счет взаимодействия элементов их вторичной структуры (рис. 16Б).
В физиологических условиях однотяжевые РНК характеризуются компактной и упорядоченной третичной структурой, которая возникает за счет взаимодействия элементов их вторичной структуры (рис. 16Б).
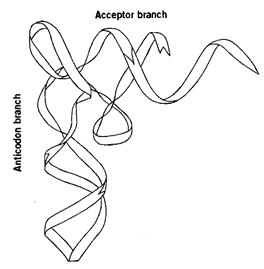
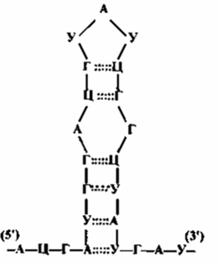 Рис. 16. Структура РНК:
Рис. 16. Структура РНК:
А – участок вторичной структуры в цепи РНК; Б – схема третичной структуры тРНК
Виды РНК
Матричная, или информационная, РНК (мРНК) переносит генетическую информацию из клеточного ядра в цитоплазму.Составляет около 5% всей клеточной РНК. Зрелые мРНК имеют сходный план строения. Линейная цепь мРНК содержит несколько областей с различной функциональной ролью (рис. 17).
|
|

Рис. 17. Составные части зрелой мРНК
1. «Колпачок» (кэп) – нуклеотидная последовательность на 5′-конце мРНК, состоящая из одного-четырех модифицированных нуклеотидов. Такая структура защищает 5′-конец мРНК от действия экзонуклеаз.
2. 5′-нетранслируемый участок – последовательность из нескольких десятков нуклеотидов, которые комплементарны нуклеотидам рРНК, входящим в состав малой субъединицы рибосомы. За счет этого 5′-нетранслируемый участок выполняет функцию первичного связывания мРНК с рибосомой, но сам не транслируется.
3. Инициирующий кодон – кодон, с которого начинается трансляция мРНК. Во всех мРНК он один и тот же – АУГ (кодирует метионин).
4. Кодирующая часть содержит информацию о последовательности аминокислот в белке. В зрелой мРНК она лишена интронов – вставочных некодирующих последовательностей, т. е. имеется непрерывная последовательность смысловых кодонов, которая должна считываться в направлении 5′®3′.
5. Терминирующий кодон – один из трех «бессмысленных» кодонов: УАА, УАГ или УГА.
6. 3′-нетранслируемый участок содержит последовательности нуклеотидов, которые называют элементами нестабильности мРНК. Определенные белки клетки узнают эти последовательности, связываются с ними и стабилизируют мРНК.
7. Поли (А)-фрагмент содержит от 50 до 400 адениловых нуклеотидов. Эти фрагменты отсутствуют в молекулах гистоновых мРНК. Полагают, что поли (А)-фрагмент имеет отношение к регуляции продолжительности жизни мРНК. Согласно одной из гипотез, после того как очередная рибосома заканчивает трансляцию мРНК, от поли (А)-фрагмента отщепляются 10-15 нуклеотидов. Когда данный фрагмент исчерпывается, начинает разрушаться кодирующая часть мРНК (если отсутствует 3′-нетранслируемый участок).
Общее количество нуклеотидов в мРНК составляет обычно несколько тысяч. При этом на кодирующую часть приходится лишь 60-70% нуклеотидов.
 Рис. 18. Структура тРНК (на примере тРНКPhe)
Рис. 18. Структура тРНК (на примере тРНКPhe)
|
 Транспортные РНК (тРНК) – короткие молекулы (70-90 нуклеотидов), главной функцией которых является присоединение аминокислот и перенос их в белоксинтезирующий аппарат клетки. Количество различных тРНК в клетке – несколько десятков: от одного до шести видов для каждой из 20 аминокислот. Виды тРНК, способные связывать одну и ту же аминокислоту, называются изоакцепторными. Специфичность тРНК обознается верхним индексом, например тРНКАла.
Транспортные РНК (тРНК) – короткие молекулы (70-90 нуклеотидов), главной функцией которых является присоединение аминокислот и перенос их в белоксинтезирующий аппарат клетки. Количество различных тРНК в клетке – несколько десятков: от одного до шести видов для каждой из 20 аминокислот. Виды тРНК, способные связывать одну и ту же аминокислоту, называются изоакцепторными. Специфичность тРНК обознается верхним индексом, например тРНКАла.
Среди нуклеотидов тРНК высоко содержание минорных нуклеотидов (около 10%). Благодаря образованию участков вторичной структуры, цепь тРНК имеет характерную форму «клеверного листа» (рис. 18).
В этой структуре четыре двухцепочечных и пять одноцепочечных участков. Поскольку минорные нуклеотиды, как правило, не способны к комплементарным взаимодействиям, они содержатся в основном в одноцепочечных локусах.
Для одноцепочечных участков приняты следующие названия:
· акцепторная ветвь – участок на 3′-конце тРНК из четырех нуклеотидов, место присоединения аминокислоты;
· антикодоновая петля – участок из 7 нуклеотидов в середине цепи; три из них выполняют функцию антикодона, который комплементарно взаимодействует с соответствующим кодоном мРНК;
· дигидроуридиловая, псевдоуридиловая и не всегда имеющаяся добавочная петли способствуют формированию специфичной для данной тРНК третичной структуры.
Особенность тРНК – наличие стабильной третичной структуры. Четыре двухцепочечных участка, попарно сближаясь, образуют примерно два витка двойной спирали, расположенные почти перпендикулярно друг другу – так, что молекула приобретает Г-образную форму (рис. 16Б).
 Рибосомные РНК являются структурной основой для формирования субъединиц рибосом. Среди азотистых оснований в рРНК выше, чем обычно, содержание гуанина и цитозина. Встречаются также минорные азотистые основания, но не столь часто, как в тРНК – примерно 1%. Во вторичной структуре рРНК много двухцепочечных участков и петель (рис. 19).
Рибосомные РНК являются структурной основой для формирования субъединиц рибосом. Среди азотистых оснований в рРНК выше, чем обычно, содержание гуанина и цитозина. Встречаются также минорные азотистые основания, но не столь часто, как в тРНК – примерно 1%. Во вторичной структуре рРНК много двухцепочечных участков и петель (рис. 19).
Рис. 19. Структура 16S-рРНК прокариотической и 18S-рРНК эукариотической клеток
 Рибосомные РНК, а также образуемые ими субъединицы рибосом принято обозначать по их константе седиментации (S). Седиментация (от лат. sedimentum – осадок) – оседание частиц, взвешенных в жидкости, при центрифугировании. Под действием центробежных сил, намного превосходящих силу тяжести, даже сравнительно небольшие макромолекулы, такие как тРНК, разделяются и распределяются в строгом соответствии со своими размерами. Измерение коэффициента седиментации макромолекулярных комплексов обычно используют для определения их общей массы и количества входящих в их состав субъединиц.
Рибосомные РНК, а также образуемые ими субъединицы рибосом принято обозначать по их константе седиментации (S). Седиментация (от лат. sedimentum – осадок) – оседание частиц, взвешенных в жидкости, при центрифугировании. Под действием центробежных сил, намного превосходящих силу тяжести, даже сравнительно небольшие макромолекулы, такие как тРНК, разделяются и распределяются в строгом соответствии со своими размерами. Измерение коэффициента седиментации макромолекулярных комплексов обычно используют для определения их общей массы и количества входящих в их состав субъединиц.
Рис. 20. Состав цитоплазматической рибосомы эукариот (в скобках указано
количество нуклеотидов в цепи рРНК)
Размеры молекул рРНК в рибосомных субъединицах у разных организмов варьируют (рис. 20), но сложная ее структура остается относительно постоянной. Кроме рРНК в состав рибосомы входит значительное число белков, но многие из них, по-видимому, не являются необходимыми для функционирования рибосом. Можно предположить, что именно молекулы РНК, а не белковые молекулы катализируют многие реакции, протекающие на рибосомах. Рибосомные белки при этом лишь усиливают функции рРНК.
Функции РНК
РНК многофункциональна. К ее основным функциям относятся:
1. Передача наследственной информации о структуре белка от ДНК к белоксинтезирующему аппарату клеток (мРНК).
2. Формирование структуры рибосом (рРНК).
3. Специфическое связывание и перенос аминокислот к рибосомам (тРНК).
Вместе с тем РНК выполняет ряд других функций:
· участвует в репликации ДНК, выступая в роли затравок (праймеров), необходимых для инициации синтеза комплементарных цепей ДНК;
· является матричной молекулой в процессе обратной транскрипции (биосинтезе ДНК на матрице РНК) и при собственной репликации у РНК-содержащих вирусов и фагов; матричные свойства РНК реализуются также в процессе наращивания теломерных повторов в молекулах ДНК;
· осуществляет специфический катализ химических реакций в клетке.
В 1982-1983 гг. в лабораториях Т. Чека и С. Олтмана (США) было сделано сенсационное открытие, осуществившее революцию в биохимии и молекулярной биологии: показано, что РНК может быть специфическим катализатором биохимических реакций. В течение всей предшествующей истории биохимии на протяжении десятилетий утверждалось, что биохимический катализ – функция исключительно белков-ферментов. По аналогии с белками-ферментами – энзимами – каталитические РНК были названы рибозимами. В настоящее время рибосому принято рассматривать как рибозим. Все имеющиеся экспериментальные данные свидетельствуют о том, что синтез полипептидной цепи белка в рибосоме катализируется рибосомной РНК, а не рибосомными белками.
Концепция «Мир РНК»
Открытие многофункциональности РНК, ее каталитических функций привело к созданию концепции «Мир РНК» – концепции мира, который, вероятно, возник и существовал задолго до оформления ныне существующего «ДНК-белкового мира». По ряду причин именно РНК, а не ДНК могла представлять собой первичный генетический материал:
· во-первых, и в химическом синтезе, и в биохимических реакциях рибонуклеотиды предшествуют дезоксирибонуклеотидам; дезоксирибонуклеотиды – продукты модификации рибонуклеотидов;
· во-вторых, в самых древних, универсальных процессах жизненного метаболизма широко представлены именно рибонуклеотиды, а не дезоксирибонуклеотиды, включая основные энергетические носители (АТФ и т. п.);
· в-третьих, репликация РНК может происходить без какого бы то ни было участия ДНК, а механизм редупликации ДНК даже в современном живом мире требует обязательного участия РНК-затравки в инициации синтеза цепи ДНК;
· в-четвертых, обладая всеми теми же матричными и генетическими функциями, что и ДНК, РНК способна также к выполнению ряда функций, присущих белкам, включая катализ химических реакций.
Таким образом, имеются все основания рассматривать ДНК как более позднее эволюционное приобретение – модификацию РНК, специализированную для выполнения функции воспроизведения и хранения уникальных копий генов в составе клеточного генома без непосредственного участия в биосинтезе белков.
|
из
5.00
|
Обсуждение в статье: Полиморфизм двойной спирали ДНК |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы