 |
Главная |


Особенности реализации наследственной информации у эукариот
|
из
5.00
|
Геном эукариот организован сложнее, чем у прокариот. Для него характерен хромосомный уровень организации. В хромосомах ДНК находится в окружении белков. В геноме эукариот имеется много избыточной ДНК. В генетическом материале эукариот находятся неинформативные участки – интроны, которые между между информативными - экзонами. Лнтроннс-экзонная организация генов у эукариот определяет необходимость преобразования первичного транскрилта (пре-информационной РНК'- продукта транскрипции) в зрелую и-РНК. Она долина быть освобождена от неинформативных участков и защищена против разрушающего воздействия ферментов цитоплазмы.
Кроме того, у эукариот появляется ядерная мембрана, которая чространственно разобщаем место хранения генетической информации (хромосомы, находящиеся в ядре) и место синтеза пептидной цепи (рибосомы, находящиеся в цитоплазме). Иными словами, у эукариот процессе транскрипции и траслятши оказываются разобщенными как пространственно (ядерной обо.. 1кой), так и во времени (процессами созревания и-РНК).
Таким образом в ходе реализации наследственной информации' у эукариот могло выделить следующие этапы:
а) транскрипция
б) посттранскрипционные процессы (процесскнг)
в) трансляция
г) посттрансляционные процессы. <*
"а" и "б" протекают в ядре, "в" и "г" протекают в цитоплазме.
Транскрипция - процесс.переписывания информации, зашифрованной в молекуле дНК на молекулу и-РНК - осуществляется при участии фермента РНК-полимеразы. Этот фермент катализирует оборку И--РИК в направлении от 5' к 3* концу. Транскрипция осуществляется в соответствии с принципами комплементарности и антилараллеяькости. Вот почему она мо&ет происходить на одной из двух полпнуклеотидных цепей дНК, а именно, на той, которая начинается с Зг конца, с"; а цепь называется кодогенной.

транскрипция иРНК
кодогенная (матричная)цепь ДНК
структурная часть гена

В участке у молекулу аНК, соответствущем отдельному гену, перед структурной часть©, в которой зашифрована последовательнооть "аминоквслот. в--пептиде, осязательно располагается последователь-:юсть нуклеотидов, узнаваемая РПК-полимеразой. Такая последовательность называется промотором.
РНК-пслимераза находит промотор, взаимодействует с ним и после зтого, двигаясь вдоль молекулы дКК, обеспечивает постепенную сборку молекулы и-РНК в соответствии с принципами комп-лементарности и антипараллельности. В конце структурной части гена расположен участок с особой последовательностью нуклеотидов -»те-рмилатор» Он обязательно включает один из нонсенс-триплетов ^.нв кодирующих аминокислоты.
В результате транскрипции синтезируется молекула пре-ин-формационной РНК.
Посттраяскршплонные пропеосы (птюцессинг) - это превращения, происходящие с первичным траыскриптом, направленные на образование зрелой, стабилизированной и-РНК, способной выполнять функцию матрицы при тг^сяяции, и защищенной от рагрушащвго воздействия специфических ферментов цитоплазмы.
Основные стадии щхщессинта:
а) отщепление концевых участков первичного транскрипт^:
б) формирование на 5' конце колпачка, состоящего из особой последовательности нуклеотидов;
в) формирование на 3* конце полиадениловой последовательности нуклеотидов А А А А ;
г) метилирование некоторых внутренних азотистых оснований в транскрипте, стабилизирующее молекулу РНК;
д) вырезание неинформативных участков, соответствующих интронам дНК и сшивание (сплайсинг) участков, соответствующих экзокам
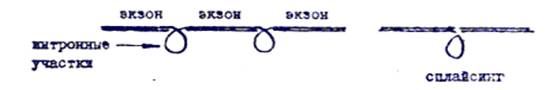
В результате процессинга у эук*>риот образуется зрелая и-РНК, характеризующаяся следующими особенностями строения:

Колпачок - особая последовательность нуклеотидов с метилированными основаниями, которая обеспечивает узнавание малых субъедгошц рибосом.
Лидер - вводная последовательность нуклеотидов, комплементарная последовательности в молекуле р-РНК малой субъединиц:; рибосомы, которая обеопечивает прикрепление и-РЙК к малой субъединице.
Стартовый кодон - триплет нуклеотидов, кодирующий в большинстве случаев аминокислоту формилметионин (АУЛ.
Кодирующая часть - последовательность кодонов, шифрутщих определенную последовательность аминокислот в соответствующей пептидной цепочке.
Трейлер - концевая часть молекулы и-РНК, включающая нок-сенс-кодон и поли-А последовательность.
Трансляция- процесс сборки пептидной цепи, происходящий в цитоплазме на рибосомах на основании программы, содержащейся в и-РНК.
8 467k 27 *
Основные сазн тргнслятзгл: инициация
элонгация
терминация Инициация трансляции предполагает ел едущие события:
а) с помогая колпачка и-РНК находит в цитоплазме малую субъеда-
НЕПУ рибОСОМЫ,
б) с помощью лкдерной последовательности устанавливается связь „ с комплементарным участком определенно!5 фракции р-РНК и
и-РНК прикрепляется к \:алой субъеднннце, ») к стартовому кодону (АУТ) присоединяется т-РНК, несущая
формилиетгокин, р) малая субъедикица ассоциируется с большой субъединицей,в «й
ноацильном центре (АЦ) которой располагается формилметшнин.
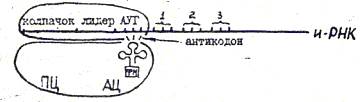
Таким образом фаза инициации завершается формированием комплекса и-РНК и рибосомы и подстановкой начальной для всех пептидных цепей аминокислоты - формилметионина.
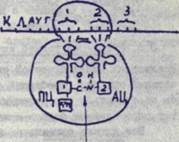
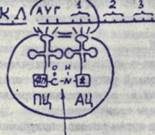 Раза элонгации, т.е. нарастание пептидной цепи, осуществляется путем постепенной подстановки аминокислот в соответствии с очередным ко доном и-РНК, который встает против аминоацильного центра.
Раза элонгации, т.е. нарастание пептидной цепи, осуществляется путем постепенной подстановки аминокислот в соответствии с очередным ко доном и-РНК, который встает против аминоацильного центра.
К этому кодону присоединяется соответствущая т-РНК, имещая комплементарный ему антикодон. Она несет определенную аминокислоту, которая располагается в аминоадкльном центре (АЦ), Т-РНК, соединенная с предыдущим ко доном, оказывается в пеп-тидильяом центре (ГЩ), где располагает свою аминокислоту (цепочку аминокислот). Между двумя аминокислотами, расположенны-ми в пептидильноы и аминоадкльном центре, при участии имеющихся здесь ферментов возникает пептидная связь -с.-//- • После установления пептидной
пептидная связь связи предыдущая т-РНК отделяется от своей аминокислоты и своего кодона и уходит в цитоплазму» а последующая т-РНК, нагруженная цепочкой аминокислот, переходит в ВД, заставляя и-РНК перемещаться вдоль рибосомы и устанавливать новый кодон против АЦ.
После прохождения через рибосому всей кодирующей части и-РНК на рибосоме собирается пептидная цепь с определенной последовательностью аминокислот.
Фаза термикацид наступает, когда в контакт с рибосомой приходит концевой участок и-РНК, который включает нонсенс-триплет, не ко-дируший никакой аминокислоты. На этом сборка пептидной цепи заканчивается.По мере освобождения 5» пептидная связь конца и-РЖ, колпачок может находить новые малые субъедини цы рибосом и пу f,ecc трансляции может повторно осуществляться на новых рибосомах. Комплекс рибосом, находящихся в контакте с одной молекулой и-РНК и синтезирующих одинаковые пептидные цепи, называется полирибосомой (по-лисомой).
Посттрансляционные процессы
В ходе предыдущих этапов реализации наследственной информации обеспечивается синтез пептидной цепа, котбрая в большинстве случаев начинается с аминокислоты формияметЕон.;с; и соответствует первичной структуре белковой молекулы. Последующие события заключаются в отщеплении форыилметионинс. в некоторых случаях осуществляется моди^Ецировакие пептида после трансляции, формируется вторичная и третичная структура белка. Иногда для некоторых белков, характеризующихся четвертичной структурой, осуществляется объединение одинаковых,либо различных лептидных цепей с образованием активно функционгрущего белка.
В зависимости от того, каковы функции белка (фермент, строительной материал, антитело и т.д.), он принимает участие в обеспечении морфо--функциональ1шх особенностей клетки (ojv^
ганжзма), т.е. в формировании определенных сложных признаков.
Это является завершающим этапом процесса реализации генетической информации.
3.5. Регуляция генной активности
Реализация наследственной информации в живых системах - это сложный процесс, требующий очень тонкой регуляции #*я того, что-'бы обеспечить в определенных клетках в -определенное время синтез определенных белков а необходимом количестве.
Все клетки организма, возникая путем митоза, получают полноценный набор генетической информации, образуемый при оплодотворении родительских гамет. Нес- ыотря на это, они отличаются по своим морфологическим, биохимическим и функциональным свойствам друг от друга. В основе этих различий лежит активное функционирование в разных клетках разных частей генома.
Большая часть генома в клетках opi-анизма находится в неактивном состоянии - репрессивном состоянии, и только приблизительно 1055 генов ^репрессированы. т.е. активно транскрибируются. Спектр транскрибируемых генов зависит от тканевой принадлежности клетки, от периода ее жизнедеятельности и периода индивидуального развития организма.
Регуляция активности генов может осуществляться на всех этапах реализации генетической информации, но наиболее экономически выгодной является регуляция на стадии транскрипции.
Основная масса генов, активно функциснирующих в большинстве клеток организма на протяжении онтогенеза, - это гены, которые обеспечивают синтез белков общего назначения (белки рибосом, хромосом, мембран я т.д.), т-ГЯК и р-РНК. Транскрибирование этих структурных генов обеспечивается соединением РНК-полимеразы с их промоторами и не подчиняется каким-либо другим регулирующим воздействиям. Такие гены называются конститутивными, другая группа структурных генов, обеспечивающих синтез некоторых белков-ферментов, в своем функционировании зависит от различных регулирующих факторов и называетеп регулируемыми генами. Их активное функционирование, скорость и продолжительность транскрибирования могут регулироваться как генетическими факторами, так ж факторами негенетической природы. - . Генетическими факторами регуляции тг*шскридцни генов явля-
ются гены - регуляторы и операторяг Гены-регуляторы определяют синтез ^.яков-регуляторов, способных в активном состоянии соединяться с оператором, включающим или выключающим транскрипцию структурных генов. В зависимости от свойств белка-регулятора различают негативный и позитивный контроль транскрипции со стороны гена-регулятора. При негативном контроле белок-регулятор, соединяясь с оператором, прекращает (выключает) транскрипцию. Такой белок называется репрессором. При позитивном контроле белок-регулятор, соединяясь с оператором, включает транскрипцию. В таком случае продукт гена-регулятора называется апоиндуктором.
| Вид генетического контроля | Вид белка-регулятора | Исходная форма белка-регулятора | Экспрессия гена (транскрипция) |
| Негативный контроль | Репрессор Репрессор | Активный Неактивный | - + |
| Позитивный контроль | Апоиндуктор Апоиндуктор | Активный Неактивный | + - |
Таким образом наряду со структурными генами в геноме имеются гены-регуляторы, которые, обеспечивая репрессию или дерепрессию структурных генов, регулируют процессы синтеза в клетке.
Наряду о генетическими факторами в регуляции экспрессии генов важная роль принадлежит факторам негенетической природы - эффекторам. К ним относятся вещества небелковой природы, расщепляемые или синтезируемые в клетке при участии разд-^ых ферментов.
| Вид эффектора | Вид белка-регулятора | Исходная форма белка-регулятора | Характер взаимодействия эффектора с белком-регулятором | Экспрессия генов (транскрипция) |
| Индуктор Индуктор | Репрессор Апоиндуктор | Активный Неактивный | Инактивация Активация | + + |
| Корепрессор Корепрессор | Репрессор Апоиндуктор | Неактивный Активный | Активация Инактивация | - - |
В аавксжмостн от того, как эффектор воздействует на активность генов, различают индукторы,включапзие транскрипцию генов, и ко-репрессоры. выключающие ее. действие эффектора заключается в его взаимодействии с белком-регулятором, при котором он либо активируется и может соединяться с оператором, либо инактиви1:•? этся в теряет способность соединяться с оператором.
Таким образом экспрессия генов является результатом регулирующего воздействия на процессы транскрипции как со стороны самсго генома (гены - регу, тторы и операторы), так и со стороны факторов вегеяетической природы.
Регуляция транскрипции у прокариот
Езучениб регуляции экспрессии генов на стадии транскрипции у прокариот привело в созданию в 1961 г. модели оперона (1акоб и Моно).
Оперся - это тесно связанная последовательность структурных генов, определяющих синтез группы ферментов д*." ,<акой-либо одной цепи биохимических реакций и регулирующаяся как едино„ целое.
Модель оперона структурные гены
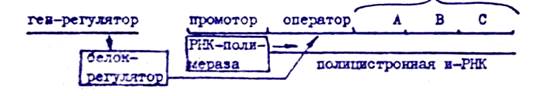
Особенностью прокариот является транскрибирование и-РНК со всех структурных генов оперона. Такал полицистронная и-РНК в дальнейшем разрезается на фрагменты, соответствующие матрицам для синтеза отдельных ферментов. Цепи структурных генов оперона всегда предшествует промотор, узнаваемый РНК-полимеразой. 7 конститутивных гонов этого достаточно для осуществления транскрипции. У регулируемых генов между промотором и структурнши генами располагается оператор - последовательность нуклеотидов, которая узнается белком-регулятором, находящимся в активном состоянии. Пример функциовдрования , актозного оперона 6..Сое/
При отсутствии в среде лактозы активнг" репрессор, взаимодействуя с оператором, репрессирует гены ABC - транскрипции нет. Появление в среде лактозы инактивирует репрессор, он не соединяется с оператором, и осуществляется транскрипция генов ABC,
отвечающих за синтез ферментов, которые расщепляют лактозу.
Пример негативного контроля функции лактозного оперона у E.coli
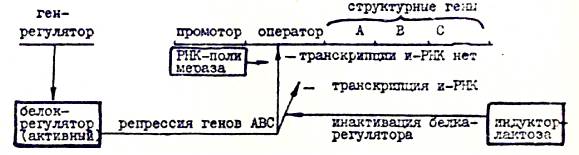
Уменьшение содержания лактозы в результате ее ферментативного расщепления приводит к соединению активного репрессора с оператором и выключению транскриЕции генов АЗС. Особенности регуляции транскрипции у эукариот
°У эукариот оперонная организация генов не установлена. Гены, определяющие синтез 'ферментов, кателизиружих разные звенья в цепи биохимических реакций, могут быть рассеяны в геноме, и, возможно, не имеют/как у прокариот, един^ регулирующей системы (г? -^г.лятор, промотор, оператор). Ь настоящее время механизмы регуляции и координафя активности таких генов окончательно не выяснены. Однако их функционирование несомненно подчиняется регуляторным воздействиям как внутри клетки (гены-регуляторы), так и на уровне организма (гормона).
Помимо регуляции экспрессии генов на стадии тралс:ср:гп::ии, она может осуществляться и при процессинге (обсуждается роль нитронов) и в ходе трансляции и поеттрасляционнылг модификаций белков.
Несмотря на то, что регуляция на поздних этапах реализации наследственной информации экономически менее выгодна клеткам, она обеспечивает наиболее быстрый ответ на воздействие регулирующих факторов. Например прекращение трансляки/ пептидной цепи сразу дает эффект по сравнению с прекращение'.! транскрипции соответствующего гена, так. как синтезированные молекулы и-РНК еще некоторое время после окончания транзхряппда обеспечивают в цитоплазме сборку пептидной цели. В совокупности все механизмы регуляции генной активности, обеспечивает производство бглков в необходимом и достаточном в данный момент количестве.
|
из
5.00
|
Обсуждение в статье: Особенности реализации наследственной информации у эукариот |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы