 |
Главная |


Динамика развития элементов ушной раковины в зависимости от возраста и размеров эмбриона
|
из
5.00
|
| Возраст | Размеры, мм | Характеристика элементов |
| IV неделя | Хорошо выражены жаберные дуги и жаберные щели. | |
| Начало V | Хорошо видны жаберные дуги и жаберные щели, особенно первые три. | |
| недели | ||
| V неделя | Хорошо видны жаберные дуги и жаберные щели. Происходит разделение мандибулярной дуги на верхнечелюстной и нижнечелюстной отростки. Позади гиоидной дуги хорошо виден слуховой пузырь. | |
| V неделя | По краям первой жаберной щели начинают образовываться небольшие бугорки, которые в дальнейшем дадут начало наружному уху. | |
| Конец V недели | Жаберные дуги плохо заметны. Из жаберных щелей сохранилась только первая,по заднему краю которой формируются бугорки наружного уха. Остальные жаберные щели закрыты кожным выростом второй жаберной дуги. | |
| Начало VI недели | Эмбрион приобретает черты человека. Хорошо виден широкий наружный слуховой проход, ограниченный двумя складками кожи, которые лишь слегка выступают над поверхностью. | |
| VII неделя | Размеры наружного слухового прохода уменьшаются. Отчетливо видна ушная раковина, но размеры ее небольшие. | |
| Начало VIII недели | Ушная раковина увеличивается и начинает приобретать дифференцированную форму. | |
| VIII неделя | Хорошо развита ушная раковина, принимающая типичную форму. | |
| Х неделя | Ушная раковина отчетливо выражена. Завиток почти закрывает отверстие слухового прохода. Имеются элементы корня завитка и козелок. | |
| XI неделя | Ушная раковина сформирована во всех своих частях. Хорошо видны завиток и проти- | |
| XVIII неделя | возавиток, козелок и противокозелок, корень завитка. Хуже оформлена конха. Ушная раковина хорошо развита и отличается от уха человека лишь своими меньшими размерами. |
Они располагаются позади зачатков глаз, которые хорошо различимы в виде небольших круглых пятнышек (рис. 5).


Рис. 5. Основные моменты формирования жаберного аппарата в первые два месяца эмбриогенеза у зародыша человека (по Л. И. Фалину, 1976): а— на 4-й неделе, при длине 6мм; б—в конце 5-й недели, при длине 14 мм; в — на 6-ой неделе, при длине 18 мм.
Обозначения:
1 — первая жаберная дуга (мандибулярная);
2 — вторая жаберная дуга (гиоидная);
3 —-третья жаберная дуга;
4 — четвертая жаберная дуга;
5 — бугорки жаберных дуг, формирующие элементы ушной раковины;
6 — жаберная щель, формирующая наружный слуховой проход.
У зародышей длиной 11—14 мм в середине^пятой и в начале шестой недели по краю первой жаберной щели начинают формироваться небольшие бугорки, которые в дальнейшем дают начало наружному уху.
К концу второго месяца зародыш уже приобретает черты человека и достигает длины до 25—30 мм. Шейный изгиб сглаживается, голова приподнимается; нормализуются соотношения отдельных структур лица, за исключением ушных раковин, которые располагаются очень низко от области лица — на шее. К этому периоду уже завершается основное дифференцирование нервной трубки на будущие отделы мозга. Начало этого процесса закладывается в шейной области и распространяется в каудальном и краниальном направлениях. Образовавшиеся спинальные ганглии начинают формировать периферические чувствительные нервные волокна., которые направляются в соответствующие сегменты тела. В этот период эмбрион как раз уже завершает сомитную стадию развития, то есть стадию сегментации (Л. И. Фалин, 1976) и периферические чувствительные нервы распределяются на основе сегментарного принципа.
Расположение жаберных элементов и зачатков будущего наружного уха на шее. приводит к тому, что чувствительная соматическая иннервация кожных покровов уха формируется за счет нервных ветвей шейных нервов, то есть будущего шейного сплетения. Именно этим можно объяснить, почему в иннервации значительной части уха, которое у человека расположено в области головы, участвуют нервы шейного сплетения (Сд—С^}. Однако эти нервы не иннервируют все области уха в связи с тем, что часть элементов уха закладывается в эмбриогенезе не только из кожных образований жаберного аппарата, но из других его элементов (рис. 6). Жаберный аппарат, который у эмбриона человека является лишь кратковременным, мимолетным эпизодом онтогенеза, играет огромную роль в физиологии животных, обитающих в водной среде. Для обеспечения нормальной работы этой жизненно важной функциональной системы требуется участие ряда структур мозга и соответствующие нервные связи (см. ниже).
Итак, наружное ухо человека формируется из жаберного аппарата и в этом процессе участвует главным образом первая жаберная щель и ее окружающее утолщение.
Рассмотрим эмбриогенез жаберного аппарата и его производных у человеческого зародыша. Местом образования жаберного аппарата является передний — оральный отдел кишки. Уже это обстоятельство объясняет, почему в иннервации жаберного аппарата участвует блуждающий нерв (п. vagus), однако участие блуждающего нерва объясняется не только этим обстоятельством, а другими более важными, о чем будет связано при рассмотрении филогенеза.
Жаберный аппарат (branchial apparatus) состоит обычно из пяти пар жаберных карманов и такого же числа жаберных дуг и щелей. Последняя, пятая, пара жаберных карманов в эмбриогенезе человека является рудиментарным образованием. В процессе эмбриогенеза первыми образуются жаберныг карманы'в виде выпячивания эндодермы в область боковых стенок глоточного или жаберного отдела передней кишки. Жаберный аппарат самым тесным образом' связан с глоткой и началом кишечной трубки. Поэтому нет ничего неожиданного в том, что в иннервации отдельных структур жаберного аппарата принимают участие нервы, иннервирующие глотку и кишечную трубку, в частности блуждающий (X) и языко-глоточный (IX) нервы. Навстречу эндодермальным выступам, образующим жаберные карманы, имеются эктодермальные выпячивания, образующие жаберные щели в области шеи. Однако у человеческого зародыша эти щели полностью не прорываются в глоточную полость и'поэтому настоящие жаберные щели, характерные для водных позвоночных (рыбы, хвостатые амфибии, личинки бесхвостых амфибий) не образуются. Участки мезенхимы, заложенные между соседними жаберными карманами и щелями, разрастаясь, образуют на передне-боковой поверхности шеи валикообразные утолщения — жаберные дуги (рис. 6).
В мезенхимальной основе жаберных дуг, которые отделены друг от друга жаберными щелями, проникают сосуды (аортальные дуги) и нервы; развиваются мышцы и хрящевой скелет.
Самой крупной является первая жаберная дуга (рис. 6), называемая мандибулярной (нижнечелюстной), из которой далее формируются элементы челюстей. Вторая жаберная дуга называется гиоидной и дает начало подъязычной коcm..(os. hyoideus).
Третья дуга идет частично на образование щитовидного Хряща. Ниже расположенные четвертая и пятая жаберные дуги значительно меньшего размера. Они постепенно •редуцируются, срастаются с расположенными выше дугами. Кожная складка, которая начинает расти от нижнего края второй жаберной дуги, постепенно закрывает нижние жаберные дуги и образует глубокий синус на шее (sinus cervicalis).'3TOTсинус вначале сообщается с внешней поверхностью шеи при помощи небольшого отверстия,которое быстро зарастает. При дефектах эмбрионального развития на шее ребенка может быть фистулёзный ход. сообщающийся иногда с полостью глотки. 
V VII IX ХС,-С, :
Рис. 6. Эмбриогенез ушной раковины: а—участки ушной раковины, формируемые из соответствующих жаберных бугорков (по R.Воиг-diol, 1975); б— иннервация жаберных дуг и жаберных щелей; в—вертикальный разрез через первую жаберную щель.
Видны: М — первая (мандибулярная) и Н — вторая (гиоидная) Дуги.1—6 — бугорки жаберных дуг, из которых формируется ушная раковина.
Штриховкой обозначены участки иннервации соответствующиминервами.
Из жаберных карманов постепенно формируются: полость среднего уха и евстахиева труба (первая пара), миндалины (вторая пара), околощитовидные железы и тимус (третья и четвертая пара). Из жаберных дуг возникают также зачатки языка и щитовидной железы.
После образования шейного синуса путем заращения кожной складки от второй жаберной дуги на шее остается заметной лишь первая жаберная щель и окружающие ее первая и вторая жаберные дуги.
В процессе эмбриогенеза первая жаберная щель превращается в наружный слуховой проход, а из кожной складки, окружающей наружный слуховой проход, то есть из области первой и второй жаберных дуг, прилежащих непосредственно к жаберной щели, развивается ушная раковина. Так как первая жаберная дуга — мандибулярная дает начало верхней и нижней челюсти, то нет ничего неожиданного в том, что в ее иннервации участвуют мощные ветви тройничного нерва. В связи с этим становится понятным, почему в иннервации ушной' раковины, в частности ее переднего отдела (особенно в области козелка, начала завитка, межкозелковой области, надкозелковой области и других областей), образованного частью первой жаберной дуги, участвуют ветви тройничного нерва. Ухо как раз расположено у сустава нижней челюсти.
В образовании ушной раковины участвует и вторая жаберная дуга (рис. 6), а также ряд других трансформированных элементов жаберного аппарата. Все нервы, участвующие в иннервации отдельных структур жаберного аппарата эмбриона, в дальнейшем принимают участие в иннервации ушной раковины, образованной из элементов жаберного аппарата. Так как из жаберного аппарата образуется ряд важных органов области лица и шеи (верхняя и нижняя челюсти, язык, миндалины, евстахиева труба, среднее и наружное ухо), то в формировании его иннервации принимают участие различные нервы, которые в дальнейшем переходят на органы, образованные из элементов жаберного аппарата.
Например, язык, который развивается из нескольких зачатков, имеющих вид бугорков и расположенных на дне первичной ротовой полости, в области вентральных отделов первых трех жаберных дуг (Л. И. Фалин, 1976) получает иннервацию от нескольких нервов, принадлежащих этим жаберным дугам — тройничный (V), блуждающий (X) И ЯЗЫКО-ГЛОТОЧНБ1Й (IX).
Ветви этих же нервов иннервируют и другие органы, образованные из тех же жаберных элементов, например, наружное ухо.
Полость среднего уха возникает из первого жаберного кармана. Узкий конец этого кармана, открывающийся в глотку, дает начало слуховой или евстахиевой трубе, а широкий конец образует барабанную полость. Одновре* менно с этим происходит формирование ушной раковины и наружного слухового прохода. Они являются главным образом производными первой жаберной щели и окружающей ее мезенхимы. Вокруг входа в жаберную щель в течение второго месяца эмбрионального периода появляется группа бугорков (рис. 5), возникающих из ткани мандибулярной (первая) и гиоидной (вторая) жаберных дуг. Эти бугорки» срастаясь вместе, образуют закладку ушной раковины. В. начале третьего месяца в ее толще начинает формировать.. ся эластичный хрящ, который продолжается в наружныйслуховой проход, образуемый жаберной щелью. Согласно мнению Bourdiol (1975), вокруг жаберной щели образуется шесть бугорков, из коих три образуются из первой — мандибулярной дуги, три других—из второй—•гиоидной дуги (рис.. 6). , . . •
Из трех бугорков манднбулярной жаберной дуги обрат зуются следующие части наружного уха: козелок (первый бугорок), корень, завитка и его восходящая, часть, (второй бугорок), ножки противозавитка и вершина уха (третий бугорок). Из трех бугорков гиоидной дуги образу ютоя*. тело противозавитка и задняя часть ушной, раковины (чет.. вертый бугорок), противокозелок и хвостик завитка (пит тый бугорок), мочка уха (шестой бугорок). Согласно ло^ гике эмбриогенеза элементы ушной раковины, образован». ные из бугорков первой жаберной дуги, должны имен» афферентную иннервацию ветвями тройничного нерва» а элементы, образованные из бугорков второй жаберной дуги,—ветвями лицевого нерва, точнее— промежуточного нерва Врисберга (п. intermedtus Wrisberg;i).
Следовательно, передне-верхняя часть ушной раковины, включая козелок, 3/5 завитка, ветви противозавитка и переднюю, половину конхи, должна быть иннервирована тройничным нервом. Задне-нижняя часть, ушной раковины» содержащая мочку, хвостик завитка, тело противозавитка,. противокозелок и заднюю часть конхи, должна быть иннер' вирована промежуточным нервом. В принципе так оно,, и есть, однако с серьезными отклонениями.
Необходимо учитывать, что уже у эмбриона 'в период закладки жаберных дуг и начала их трансформации в элементы будущей ушной раковины кожа, покрывающая ету .поверхность, получает афферентную иннервацию от шейных нервов; что тройничный и лицевой нервы иннервируют не просто кожу, покрывающую жаберные дуги, а элементы, расположенные глубже,—мезодерму и эндодерму, 'имеющие функциональное значение в работе жаберного аппарата у предшествующих форм позвоночных, стадию которых проходит во время эмбриогенеза зародыш человека.
• При формировании элементов ушной раковины из жаберного аппарата в «наследство» передаются все нервы, в том числе и нервы шейного сплетения. Поэтому среди нервов, иннервирующих ушную раковину, важное место занимают ветви шейного сплетения. Согласно Bourdiol (1975), они разветвляются во всей задней половине наружной и внутренней поверхности ушной раковины (рис. 6). Тройничный нерв иннервирует только передне-верхнюю часть уха, соответствующую трехсторонней выемке, восходящей ветви завитка, ножкам противозавитка и предкозел-ковую область. Средняя часть уха, включая конху, козелок, корень завитка и тело противозгвитка, иннервируется лицевым (промежуточным) нервом. Эта точка зрения тоже не отражает истины, так как противоречит уже данным анатомии, а не эмбриологии. Мы объясняем это тем, что эмбриологические исследования проводились недостаточно тщательно, в связи с большими методическими трудностями и не были обнаружены ветви блуждающего и языко-глоточного нервов, которые непременно иннервируют жаберный аппарат и которые обнаружены среди нервов ушной раковины.
Согласно данным эмбриологии, которые приводит Bourdiol (1972, 1975), третья жаберная дуга иннервируется языко-глоточным нервом, а четвертая дуга — блуждающим нервом. Если стать на эту точку зрения, тогда следует полагать, что в формировании ушной раковины участвуют также элементы третьей и четвертой жаберных дуг, которые приносят с собой и соответствующие им нервы. Однако это противоречит достаточно убедительным данным эмбриологии (Л. И. Фалин, 1976) о формировании ушной раковины лишь из элементов первых двух жаберных дуг. Поэтому остается признать, что блуждающий и языко-глоточный нервы, несомненно, иннервируют первые две жаберные дуги, иначе откуда же они берутся на ушной раковине?— Это вполне объясняется функциональными особенностями жаберного аппарата. Соответствующие доказательства представлен» в литературе по сравнительной анатомии и физиологии позвоночных.
То, что не удается проследить в процессе онтогенеза; учитывая калейдоскопическую быстроту трансформации отдельных органов и тканей у эмбриона, проще и надежнее проследить у ряда тех взрослых животных, стадии которых проходит зародыш человека. Такой подход является совершенно оправданным, так как опирается на биологический закон рекапитуляции, который гласит, что в процессе индивидуального развития повторяются многие черты организации отдаленных предков (И. И. Шмальгаузен, 1947). Например, при эмбриональном развитии высших наземных позвоночных (включая человека) в зародышевом состоянии развиваются жаберные щели, как и у зародышей рыб, причем с теми же сосудами и нервами. Это повторение стадий развития дышавшего жабрами рыбообразного предка наземных позвоночных. Жаберный аппарат является важнейшим органом для животных, живущих в воде, причем он не ограничивается только дыхательной функцией. Для управления функцией этого сложного аппарата требуется достаточно мощная афферентная иннервация, которая обеспечивается у рыб и водных амфибий следующими краниальными нервами: V, VII, IX, X. Тройничный нерв (V) иннервирует область челюстной (мандибулярной) дуги вплоть до бризгальца, которое является редуцированной жаберной щелью. Далее, в области подъязычной (гиоидной) дуги иннервация осуществляется лицевым (промежуточным) нервом (VII). От первой жаберной щели до области второй дуги иннервация осуществляется языко-глоточным нервом (IX) и блуждающим нервом (X) .0 ветвях языко-глоточного и блуждающего нервов следует сказать подробнее.
Языко-глоточный нерв (IX) делится у рыб на три ветви—небную, преджабериую и жаберную. Преджаберная ветвь (г. praetrematicus), содержащая висцеральные афференты, иннервирует переднюю схенку первой жаберной щели, из которой у зародыша человека формируется наружный слуховой проход и конха. Жаберная ветвь (г. posttrematicus), также висцерально-афферентная, иннервирует область первой жаберной дуги, из которой формируется передкий отдел ушной раковины: козелок, корень и начало завитка.
Блуждающий нерв (X) у рыб образует два больших ствола — боковой нерв и жаберно-кишечный нерв (branchio-intestinalis). Последний иннервирует весь жаберный аппарат и внутренние органы. У каждой жаберной щели, начиная со второй, этот нерв отдает жаберные ветви, которые делятся на глоточные, преджаберные и жаберные ветви. Преджаберные ветви (г. praetrematicus) иннервируют передние стенки соответствующей жаберной щели (как и ветви языко-глоточного нерва). Жаберные ветви (г. posttrematicus) иннервируют область соответствующей жаберной дуги. Они содержат висцеральные афферентные волокна и двигательные волокна к жаберным мышцам. Продолжаясь дальше, жаберные ветви направляются вниз и образуют нервы, идущие к сердцу, кишечнику, а у наземных позвоночных иннервируют легкие и сердце (п. pneumo-gastricus). Таким образом, у рыб ветви блуждающего нерва, как и языко-глоточного, иннервируют не только задние (каудальные) жаберные дуги, но и передние (ростральные) дуги; по крайней мере, стенку первой дуги или первой жаберной щели. Поэтому, если верно, что в эмбриогенезе человеческий зародыш, повторяя стадию рыбы, образует жаберный аппарат, характерный для этого вида животного, то вполне вероятно, что иннервация жаберного аппарата человеческого эмбриона аналогична иннервации жаберного аппарата рыбы. А это в свою очередь означает, что жаберный аппарат человеческого зародыша, имея ветви блуждающего и языко-глоточного нервов в первых жаберных дугах, при образовании ушной раковины передает и эти нервные ветви.
Проблема иннервации ушной раковины достаточно сложная задача и требует серьезной разработки. Если этот вопрос анатомически еще как-то исследовался, хотя и остались спорные вопросы, то морфологически, точнее нейро-гистологически, совершенно не разработан. Не исследован спектр иннервирующих волокон, их гистохимические особенности; неизвестен характер рецепторов кожи ушной раковины, их распределение и концентрация в различных структурах уха. Очень важно узнать, какие рецепторные образования связаны с терминалями различных нервов ушной раковины, в частности соматическими и висцеральными афферентами.
А к л ю ч е ни е
На основании тщательного анализа многочисленных данных литературы по анатомии (В. П. Воробьев, 1942;
Р. Д. Синельников, 1974; Dejerine, 1914; Larsell, 195Г;
Crosby, Hamphrey, Lauer, 1962; Noback, 1967; ElHotf, 1969; Chusid, 1970; Curtis, Jacobson, Marcus, 1972; Bossy, 1959, 1970) и эмбриологии человека (Л. И. Фалин, 1978;
Bourdiol, 1972, 1975); по сравнительной анатомии позвоночных (И. И. Шмальгаузен, 1947; Е. К. Сепп, 1959); на основании данных клинических исследований (Nogrer, 1956, 1957, 1969, 1970, 1972; QuagliaSenta, 1961; Jarricot, Ming Wong, 1973; Joyeux, 1974; Sedane, 1974; Kropej, 1978;
Д. М. Табеева, Л. М. Клименко, 1976; В. Г. Вргралик, М. В. Вогралик, 1978; Э. Ф. Тыкочинская, 1979; Lu, 1975) нами была составлена серия рисунков (7,1-7,6), где при помощи штриховки изображены области иннервации наружной поверхности ушной раковины разными нервами. Без представления отдельных рисунков (7,1—7,5) 


Рис. 7.  Границы иннервации ушной раковины отдельными нервами (по данным собственных наблюдений)
Границы иннервации ушной раковины отдельными нервами (по данным собственных наблюдений)
1 — для нервов шейного сплетения (Cg—Cg); 2 — для тройничного нерва (V); З,— для лицевого нерва (VII);
4 — для языко-глоточного нерва (IX); 5 — для блуждающего нерва (X); 6 — перекрытие зон иннервации всеми пятью нервами.
для каждого нерва очень трудно уяснять границы иннервации уха этими нервами, так как на ушной раковине, как это видно на рисунках 6, 7, нет «чистых» областей, принадлежащих ветвлениям одного какого-либо нерва; большинство областей уха являются смешанными и иннервируются одновременно несколькими нервами. Особенно насыщенными в этом отношении являются такие области, как конха, трехсторонняя (ладьевидная) выемка, козелковая и предкозелковая область. Здесь представлены терминали почти всех пяти нервов, иннервирующих ушную раковину. Это хорошо видно на объединенном рисунке 7,6, хотя, сам по себе рисунок достаточно запутанный. Для того, чтобы представить себе особенности иннервации уха и связанные с этим возможности вызвать различные рефлекторные ре акции при стимуляции точек уха, разберем более подробно каждый из представленных нами рисунков и сравним их с данными других авторов, изображенными на рисунке 8.
Распределение термнналей нервов шейного сплетения на наружной поверхности ушной раковины указано на рисунке 7,1, откуда видно, что шейное сплетение иннервирует почти всю ушную раковину, за исключением области конхи, козелка и начала завитка. Шейное сплетение — это типичная соматическая афферентная система поверхностной чувствтельности, иннервирующая покровы тела, и составленная из волокон верхних шейных спинальных сегментов. Сведения об участии нервов корешка Ci в иннервации уха сомнительны, хотя Chusid (1970) такую возможность не исключает, по крайней мере, по данным этого автора, небольшая веточка от Ci направляется к ушной раковине позади мочки уха.
Участие Ci в иннервации мочки уха, как видно на рисунках 8,5 и 8,6, признают Nogier (1975) и Bourdiol (1975). Остальные авторы (Dejerine, 1914; Bossy, 1970; Kropey, 1976; Sedane, 1974) хотя и признают, что мочка уха иннервирована шейным сплетением (см. рис. 8,1—8,4), однако считают, что это нервы: второго (С2) корешка (Bossy, 1970), второго-третьего (Cg—Cg) корешков (Dejerine, 1914; Sedane, 1974) и даже третьего-четвертого (Сд—С^) корешков <Кгореу, 1976).
Все вышеуказанные авторы с различиями в отношении формирующих нервы корешков признают иннервацию нисходящей части завитка уха ветвями шейного сплетения (см. рис. 8), хотя небольшие, но несущественные расхождения между ними есть. Например, Воссу (рис. 8,2) указывает на область с верхушки уха, a Nogier (рис. 8,6)— с дарвинова бугорка.
Участие нервов С< в иннервации ушной раковины недостаточно обосновано данными анатомии (Sedane, 1970), но исключить это полностью невозможно, так как известно, что в любом нервном сплетении (шейном, плечевом и др.)

| Рис. 8. Топографические соотношения иннервации ушной раковины отдельными нервами (по данным литературы) 1. По Дежерину (Dejerine J. — 1914) 2. По Босси (Bossy J. — 1970) 3. По Седану (Sedane M. — 1974) 4. По Кропею (Kropej H. - 1976) 5. По Бурдиолю (Bourdiol R. — 1975) 6. По Ножье (Nogier P. — 1975) Условные обозначения: V — тройничный нерв; VII—лицевой нерв; IX— языко-глоточный нерв; X — блуждающий нерв; С,—Сд — нервы шейного сплетения. |
корешки вышележащих и нижележащих спинальных сегментов взаимно переплетаются (House, Pansky, 1967) и найти точно источник образования того или иного нерва очень трудно. Кроме того, как нам представляется, этот вопрос не имеет принципиального значения. Важным является не то, участвуют ли в формировании нервов ушной раковины корешки только Cg и Сд или к ним присоединяются еще и корешки С, и С4. Важным, принципиальным является: во-первых, участие нервов шейного сплетения вообще в иннервации ушной раковины; и, во-вторых, область, где разветвляются на ушной раковине терминали нервов шейного сплетения. Если по первому вопросу мнения всех авторов единодушны, то есть все признают, что ушная раковина иннервируется нервами шейного сплетения, то по второму — есть явные разногласия, что хорошо видно из рисунка 8.
Близкими к истине являются данные Sedane (1974), которая распространяет область иннервации ветвями шейного сплетения почти на все ухо (рис. 8,3), за исключением конхи, козелка, околокозел ковой области, восходящей части завитка, трехсторонней выемки и ножек противозавитка.
Все остальные авторы неоправданно резко суживают область ушной раковины, иннервированную ветвями шейного сплетения, что противоречит данным нейроанатомии (Crosby, Hamphrey, Lauer, 1962; Chusid, 1970; Р. Д. Синельников, 1978) и эмбриологии человека (Л. И. Фалин, 1976).
На рисунке 7,1 указана область иннервации наружной поверхности ушной раковины нервами шейного сплетения (Cg—Cg), составленная нами с учетом данных литературы (Chusid, 1970). За исключением конхи, начальной части завитка, козелка и околокозелковой области, трехсторонней выемки и ножек противозавитка, все остальное ухо иннервировано шейным сплетением. В этом отношении наше мнение совпадает с данными Sedane (1974).
Почему важно уточнить этот вопрос? Мы уже неодкратно подчеркивали, что точное знание иннервации ушной раковины позволяет правильно и обоснованно предсказывать функциональные особенности рефлекторных реакций, вызываемых с ушной раковины. В эмбриогенезе шейное сплетение, как и тройничный нерв, формируются одновременно и одинаково. Нисходящее или так называемое спинальное сенсорное ядро тройничного нерва располагается в шейных сегментах спинного мозга (см. рис. 10 и 11), достигая четвертого сегмента. По-существу это часть чувствительных клеток верхнего шейного отдела спинного мозга, волокна которых покидают спинной мозг не на уровне своих сегментов, а поднимаются наверх и выходят уже на уровне варолиева моста. Может быть поэтому первый шейный сегмент спинного мозга почти не дает корешков в шейное сплетение и образует очень скудный нерв?
В первые недели эмбриогенеза, когда жаберные дуги, жаберные щели и карманы уже сформированы и образуются периферические нервы, очень трудно отличить на этом участке тела эмбриона, где волокна шейного сплетения или тройничного ядра. Поэтому данные Bourdiol (1975) об иннервации жаберного аппарата человеческого эмбриона в перрые недели эмбриогенеза требуют серьезной корректировки, так как автор вообще не указывает на нервы шейного сплетения. Известно, что нервные волокна, образованные соседними нервами, при разветвлении в любой части поверхности тела никогда не образуют явных границ, а взаимно проникают, благодаря чему при поражении одного из нервов чувствительность в данном участке полностью не утрачивается. То же самое должно иметь место при иннервации жаберного аппарата у эмбриона ветвями шейного и тройничного сплетений и, следовательно, перейти далее в иннервацию ушной раковины. Поэтому приводимые разными авторами рисунки иннервации уха (см. рис. 8) с четкими границами для каждого нерва абсолютно неверны. Уже это одно говорит о том, что имеющиеся в литературе по аурикулярной рефлексотерапии взгляды об иннервации ушной раковины требуют серьезного пересмотра с учетом данных фундаментальной анатомии нервной системы человека.
На рисунках 7,1 и 7,2 указаны области ушной раковины, иннервированные ветвями шейного (С,—Сд) сплетения и тройничного (V) нерва, при сравнении которых видно, как далеко проникают взаимно терминали этих двух соматических афферентов на ушной раковине. По существу только область конхи лишена иннервации из этих двух источников. Данные рисунка 7,2 совпадают с данными других авторов (рис. 8) по иннервации уха ветвями тройничного нерва (Bossy, 1970; Nogier, 1975; Bourdiol, 1975;
Kropey, 1976). При сравнении всех рисунков серии 8 с рисунком 7.2 видно, что на всей наружной поверхности ушной раковины только нисходящая часть завитка от дарвинова бугорка до хвоста завитка и задняя часть мочки уха лишены веток тройничного нерва и имеют лишь иннервацию из шейного сплетения. Это обстоятельство дает нам право высказать ряд предположений об иннервации жаберного аппарата человеческого эмбриона в период закладки элементов ушной раковины.
Во-первых, если задняя часть ушной раковины, то есть область желобка, нисходящая часть завитка и мочка уха формируются из трех бугорков второй (гиоидной) жаберной дуги, как считает Bourdiol (1975), тогда остается признать, что кожа, покрывающая эту дугу, иннервируется ветвями шейного сплетения, хотя бы с внешней стороны, перекрываясь с ветвями лицевого нерва, который иннервирует внутреннюю сторону второй жаберной дуги, а не всю ее. Это предположение хорошо аргументируется данными Dejerine (1914) о распределении ветвей лицевого (VII промежуточного) нерва на ушной раковине (рис. 8,1).
Странно, что Bourdiol в упомянутой выше работе (рис. 8,5) совершенно отрицает иннервацию ушной раковины лицевым нервом и никак не объясняет, куда же деваются ветви лицевого нерва, указанные им в этой же' работе, при формировании ушной раковины из жаберного аппарата эмбриона?
Во-вторых, мы полагаем, что лицевойнерв у эмбриона иннервирует внутренние (медиальные) поверхности первой-и второй жаберных дуг, т, е. выход наружу жаберной щели (рис. 6).
Поэтому при формировании ушной раковины терминали лицевого нерва оказываются распределенными в области конхи и в ее ближайшем окружении (рис. 7,3).
Наш рисунок отличается от рисунка Dejerine (рис. 8,1) некоторыми особенностями. Согласно нашим данным, лицевой нерв полностью иннервирует всю трехстороннюю выемку, а не часть ее, как указывает Дежерин (рис. 8,1) и почти полностью всю мочку уха, за исключением ее задней части (рис. 7,3).
В отношении мочки уха наши данные серьезно расхо^ дятся с данными Dejerine и частично с эмбриологическими данными Bourdiol. Как и в случае с тройничными нервами (рис. 7,2 и 7,3), задний край уха (нисходящая часть завитка и задне-нижний край мочки уха) не снабжен веточками лицевого нерва. Согласно данным многих авторов, эта область иннервирована веточками шейного сплетения, что ещераз подтверждает первое предположение об участии нервов шейного сплетения в иннервации части жаберного аппарата и формирующейся ушной раковины.
Конха вместе с наружным слуховым проходом формируется из наружной трети первой жаберной щели, которая у эмбриона иннервирована лицевым (VII) нервом, а также языко-глоточным (IX) и блуждающим (X) нервами. Поэтому наличие ветвей VII, IX и Х нервов в области конхи и ближайшего окружения представляется закономерным. Но откуда берутся веточки лицевого нерва (рис. 7,3), а также языко-глоточного (ри". 7,4) и блуждающего (рис. 7,5) нервов в трехсторонней (ладьевидной) выемке, остается совершенно непонятным и требует объяснения.
Трехсторонняя выемка в смысле рефлекторных воздействий на ряд висцеральных и системных реакций организма мало чем уступает области конхи, что объясняется таким же обилием и разнообразием ее иннервации, как и области конхи. Однако это не объясняет причины такой иннервации: почему трехсторонняя выемка в процессе эмбриогенеза приобрела сходные с конхой анатомо-физиологические особенности? Не означает ли это, что трехсторонняя выемка лмеет такое же происхождение, как и конха?
Если внимательно посмотреть на рисунок уха (рис. 9), то можно отметить следующие особенности. Конха, как известно, состоит из двух полостей. Нижняя, большая полость (cavum conchae) связана с наружным слуховым проходом и как бы обрамлена хрящевыми утолщениями или валиками: спереди — козелком; сверху — корнем завитка;
сзади — концом противозавитка и противокозелком; снизу — межкозелковой выемкой. Верхняя, меньшая полость или чаша (cimbala conchae) окружена следующими хрящевыми образованиями: снизу — корнем завитка; спереди—началом восходящей части завитка; сверху — нижней ножкой противозавитка; сзади — нисходящей частью противо-завитка.
В отличие от нижней полости конхи никакого выхода в слуховой проход или какое-либо другое отверстие не имеет. В этом отношении трехсторонняя выемка весьма напоминает чашу конхи; она окружена хрящевым валиком, состоящим спереди из восходящей части завитка, снизу и сверху — из ножек противозавитка, сзади — из угла, образованного ножками противозавитка.
Кажется несколько неожиданным, что вся конха вместе с наружным слуховым проходом, которая в эмбриогенезе сформировалась из первой жаберной щели, у взрослого оказалась четко разделенной на две полости, одна из которых не имеет выхода в наружный слуховой проход. Более того, верхняя полость конхи по размерам и другим признакам несколько напоминает трехстороннюю выемку (рис. 9).
Поэтому есть достаточно оснований полагать, что трехсторонняя (ладьевидная) выемка, как и чаша конхи, обра^-зовались из той же части жаберного аппарата, из которой развивается слуховой проход и полость конхи, то есть из. первой жаберной щели, но при этом они отделились от нее за счет ее боковых выпячиваний и таким образом лишились связи с жаберной щелью. 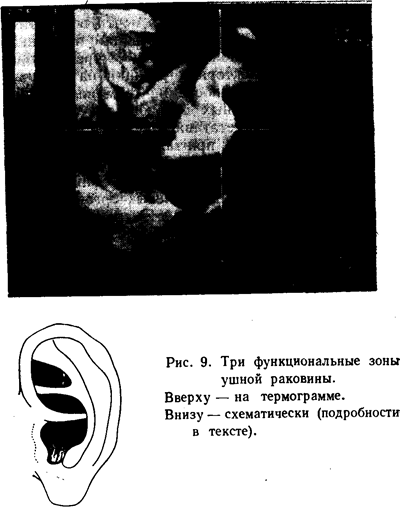
Тогда легко объяснить, почему трехсторонняя выемка имеет такую же сложную иннервацию (VII, IX, X), как и конха, со своими двумя полостями и начальной частью наружного слухового прохода. Одинаковый характер мор-фо-физиологических особенностей этих трех областей уха хорошо виден в верхней части рисунка 9, где представлена термограмма ушной раковины.
Итак, если происхождение, формирование и иннервация трехсторонней выемки аналогичны конхе, тогда есть все основания ожидать, что рефлекторные реакции, вызываемые с этой области ушной раковины, должны быть по своей природе близки к тем, которые вызываются с области конхи, и отличаются лишь по топографии своей реализации. Если конха, по мнению почти всех авторов (см. ниже),— область рефлекторных реакций на функции органов грудной и брюшной полостей, то трехсторонняя выемка — область рефлекторных реакций на функции органов малого таза, в частности на терминальные функции половых и выделительных органов, регуляция которых связана не только с вегетативной нервной системой, но и с соматической. Например, при таких формах патологии, как недержание мочи, выпадение прямой кишки и матки, спастические и атонические состояния сфинктеров прямой кишки, функции наружных половых органов и т. д. рефлекторные воздействия на область трехсторонней выемки должны быть эффективными. Наличие соматической афферентной иннервации области трехсторонней выемки веточками тройничного нерва, в отличие от области конхи где этого практически нет и имеет место лишь по. узкому краю, необходимо в связи с тем, что функции органов малого таза достаточно сильно корта кали зованы и хорошо поддаются произвольному (сознательному) контролю. Для воздействия на эти функции необходимо вовлекать в активность таламо-кортикальные проекции, что легко достигается при активации тригеминальной системы благодаря ее мощным таламо-кортикальным проекциям (Р. А. Дуринян, 1965).
Таким образом, исходя из принципов иннервации трехсторонней выемки, необходимо обратить серьезное внимание на исследование рефлекторных возможностей этой области. Опыт нашей работы, теоретический и экспериментальный анализ дают право утверждать, что область трехсторонней выемки имеет значительно меньше отношения к .проекции поверхностной чувствительности нижней крнечности (как.это ут
|
из
5.00
|
Обсуждение в статье: Динамика развития элементов ушной раковины в зависимости от возраста и размеров эмбриона |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы