 |
Главная |


Особенности различных клеточных мембран
|
из
5.00
|
Несмотря на то, что все биологические мембраны построены по единому принципу (липидный бислой с вмонтированными в него белками), их состав, физические и химические свойства различаются.
Вы, вероятно уже обратили внимание на то, что «рабочими лошадками» мембран, обеспечивающими выполнение практически всех функций, являются белки. Естественно, что в зависимости от функций той или иной мембраны, набор белков будет соответствующим. Упомянутые выше рецепторы клеточных контактов, натрий-калиевая АТФ-аза, глюкозный переносчик и т.п. расположены на плазмалемме, на мембранах митохондрий расположены другие белки, нужные для работы митохондрий, на мембранах лизосом – свои белки и т. д. При этом наблюдается следующая закономерность: чем активнее работает мембрана, тем больше в её составе белков. Внутренняя мембрана митохондрий, осуществляющих кислородный этап дыхания, содержит более 70% белков и менее 30% липидов, в то время как в плазмалемме шванновских клеток, образующих миелиновую оболочку нервов и не проявляющих высокой метаболической активности, белков менее 20% (соответственно, на долю липидов приходится более 80%).
 Различаются мембраны и составом липидного бислоя, от которого во многом зависят такие их свойства как толщина и текучесть.
Различаются мембраны и составом липидного бислоя, от которого во многом зависят такие их свойства как толщина и текучесть.
Толщина мембран определяется преимущественно длиной гидрофобных хвостов липидов. Это весьма значимый параметр: с помощью специальных белков клетка умеет отделять от мембран небольшие пузырьки, умеет она и сливать такие пузырьки с мембраной. Благодаря этим процессам может осуществляться «перетекание» мембранных пузырьков от одних органоидов к другим. Однако это слияние возможно лишь при сходной толщине липидного бислоя, поскольку гидрофильные головки более тонкой мембраны не могут контактировать с гидрофобным участком более толстой мембраны. Так плазмалемма на 2-3нм толще мембран ЭПР, и прямой контакт между ними невозможен.
От текучести/вязкости мембран зависит то, насколько эффективно они смогут выполнять защитно-барьерные функции. Очевидно, что наименее текучей мембраной должна быть плазмалемма животной клетки, лишенной жесткой поверхностной оболочки. Вязкость определяется преимущественно плотностью упаковки хвостов фосфолипидов и включением в состав бислоя дополнительных липидов с «жестко» организованным молекулярным строением.
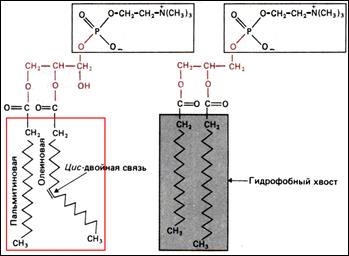 Наличие двойных связей в хвостах фосфолипидов приводит к искривлению углеродного скелета (из-за изменения угла связи) и менее плотной упаковке молекул бислоя. Мембрана становится более текучей (плазмалемма растений, внутриклеточные мембраны). Если двойные связи отсутствуют, то хвосты могут быть упакованы более плотно и текучесть мембраны снижается (плазмалемма животной клетки).
Наличие двойных связей в хвостах фосфолипидов приводит к искривлению углеродного скелета (из-за изменения угла связи) и менее плотной упаковке молекул бислоя. Мембрана становится более текучей (плазмалемма растений, внутриклеточные мембраны). Если двойные связи отсутствуют, то хвосты могут быть упакованы более плотно и текучесть мембраны снижается (плазмалемма животной клетки).
Задание.
 На верхнем рисунке изображены молекулы двух фосфолипидов, входящих в состав плазмалемм. Какой из них выделен из растительной ткани, и почему вы так считаете?
На верхнем рисунке изображены молекулы двух фосфолипидов, входящих в состав плазмалемм. Какой из них выделен из растительной ткани, и почему вы так считаете?
Как уже было сказано, снижение текучести мембран может быть достигнуто еще и включением в состав бислоя липидов с «жестко» организованным молекулярным строением. В плазмалемме животных эту роль играет холестерол (старое название – холестерин). Этот специфический липид с жесткой кольцевой структурой (рисунок справа) «связывает» соседние молекулы, снижая их подвижность. Таким образом, организм животных (а значит – и человека) нуждается в холестероле и даже производит его самостоятельно.
В мембранах митохондрий, эндоплазматического ретикулума и во многих других цитоплазматических мембранах содержится мало холестерина и сравнительно много полиненасыщенных жирных кислот. В связи с этим мембраны цитоплазмы менее жесткие, чем плазмалемма.
II. Протоплазма (цитоплазма + ядро).
Цитоплазма.
Цитоплазма – это часть живой клетки (протопласта) без плазмалеммы и ядра. В состав цитоплазмы входят: цитозоль, органоиды (включая цитоскелет) и включения (иногда включения и содержимое вакуолей к живому веществу цитоплазмы не относят).
В 1830 г. Я. Пуркинье предложил термин «протоплазма» для обозначения живого вещества в целом. Далее Р. Кёлликер (1862) ввел термин «цитоплазма» для обозначения материала, окружающего ядро. И. фон Ганштейн (1880) ввел термин «протопласт» для обозначения части клетки без оболочек, клеточного сока и включений. В настоящее время протопластом называют живую клетку, лишенную стенки.
Специализированная периферическая часть цитоплазмы в животных клетках называется эктоплазма. Здесь практически отсутствуют мембранные органоиды. В эктоплазме сосредоточены определенные элементы цитоскелета и ферментные системы трансмембранного транспорта и гликолиза. Эта часть цитоплазмы обладает повышенной вязкостью. Глубокие слои животной клетки называются эндоплазма. Здесь находится ядро и большинство органоидов клетки. Она обладает пониженной вязкостью.
Цитозоль
Цитозоль (гиалоплазма, цитоплазматический матрикс) – это основное вещество цитоплазмы. Цитозоль представляет собой водорастворимую часть цитоплазмы. Содержит около 90 % воды, в которой растворены макромолекулы и молекулярные комплексы (образующие коллоидный раствор), а также малые молекулы и ионы (образующие истинный раствор). В целом матрикс представляет собой жидкий коллоидный раствор – золь. При определенных условиях цитозоль переходит в студневидное состояние – гель. Переходы золя в гель и геля в золь – это нормальное состояние физиологически активной клетки; с этими переходами связано движение цитоплазмы, амебоидное движение клеток и изменение их формы.
Функции цитозоля: место хранения биологических молекул; среда для протекания биохимических реакций; место хранения включений; транспорт веществ; поддержание постоянства внутриклеточной среды (рН, водно-солевого режима и т.д.).
Органоиды
Одномембранные органоиды
К одномембранным органоидам относятся: эндоплазматический ретикулум, аппарат Гольджи, лизосомы, пероксисомы, сферосомы, вакуоли и некоторые другие. Все одномембранные органоиды образуют единую вакуолярную систему, которая обеспечивает разделение цитоплазмы на компартменты – отсеки, в которых протекают различные реакции.
У прокариот вакуолярная система, построенная на основе постоянных внутриклеточных мембран, отсутствует. Ее функции выполняют впячивания плазмалеммы – мезосомы.
Эндоплазматический ретикулум (ЭПР), или эндоплазматическая сеть (ЭПС)
ЭПР был открыт К. Портером в 1945г. в ходе электронномикроскопических исследований клетки.
ЭПР представляет собой систему полостей, цистерн и каналов, отгороженных от остальной цитоплазмы мембраной и соединяющихся друг с другом. Эндоплазматический ретикулум – «государство в государстве»: все его полости и цистерны непрерывно соединены, и нигде не открываются ни в цитоплазму, ни во внеклеточную среду. Более того, и перинуклеарное пространство (пространство между двумя мембранами оболочки ядра) является частью эндоплазматического ретикулума, оно также ни на одном участке не соединено ни с цитоплазмой, ни с кариоплазмой ядра. ЭПР обеспечивает функциональную взаимосвязь всех органоидов клетки между собой, объединяет все внутриклеточные мембранные структуры в единую систему, пространственно разделяет клетку. По системе каналов осуществляется транспорт веществ.
Эндоплазматический ретикулум имеется у всех эукариот и существует в виде двух типов: гранулярного (шероховатого) эндоплазматического ретикулума и агранулярного (гладкого) эндоплазматического ретикулума.
Гранулярный – грЭПР (шероховатая ЭПС) представлен системой взаимосвязанных плоских цистерн, на поверхности которых расположены рибосомы. Таким образом часть рибосом свободно плавает в цитоплазме, а другая часть «сидит» на грЭПР. Зачем нужно разделение рибосом на две группы? Дело в том, что эти рибосомы синтезируют разные белки:
- на свободно плавающих рибосомах синтезируются белки цитоплазмы, ядра и некоторые белки митохондрий и хлоропластов (остальные белки этих двух органоидов синтезируются на их собственных рибосомах);
- на рибосомах гранулярного ЭПР синтезируются белки самого ретикулума, аппарата Гольджи, лизосом, наружной мембраны и белки, секретируемые клеткой.
Белки, которые синтезируются на рибосомах гранулярного ЭПР, сразу же попадают в полость ретикулума. Хотя рибосомы расположены на наружной (т. е. цитоплазматической) стороне мембраны ЭПР, белок в процессе синтеза «проталкивается» через эту мембрану во внутреннее пространство ретикулума. Уже потом из ЭПР белки попадут в различные органоиды, войдут в состав мембран или будут секретированы, но первым этапом их внутриклеточного путешествия служит эндоплазматический ретикулум. Белки, которые должны синтезироваться на рибосомах грЭПР и попасть в полость ретикулума, содержат на N-конце особую последовательность аминокислот – лидерный пептид. Эта последовательность служит как бы специальной меткой, благодаря которой клетка «распознает», какие белки должны попасть в полость ЭПР, а какие нет. В полости грЭПР лидерный пептид отщепляется.
Агранулярный – агрЭПР (гладкая ЭПС) образован системой разветвленных трубочек.
В полости агранулярного ЭПР происходит биосинтез липидов и полисахаридов;
агрЭПР выполняет и функцию депо: например, в сократимых клетках здесь происходит накопление ионов кальция. В состоянии покоя в цитоплазме клетки ионов Са2+ содержится около 10-7М, а в полости эндоплазматического ретикулума – 2-3•10-3М. Ионы Са2+ закачиваются в полости ретикулума специальным ферментом – Са2+-АТФ-азой, естественно, за счет энергии АТФ. Ведь перекачка идет оттуда, где Са2+ мало, туда, где его много, следовательно, затраты энергии необходимы. Функция запасания Са2+ особенно важна для мышечных клеток (cм. сокращение мышцы).
В агранулярном ретикулуме печени происходит детоксикация ядовитых веществ.
Трехмерная модель и электронная микрофотография ЭПР.
 Пожалуй, одной из важнейших функций всего ЭПР является синтез клеточных мембран. Мембраны всех одномембранных клеточных органоидов (т. е. кроме мембран митохондрий и пластид), синтезируются в ЭПР. Ядерная мембрана, можно сказать, образована самим эндоплазматическим ретикулумом. К другим органоидам мембранный материал доставляется благодаря тому, что от ЭПР все время отшнуровываются небольшие мембранные пузырьки. Часть вакуолей имеет ретикулярное происхождение, то есть они образуются путем отшнуровывания от эндоплазматического ретикулума. Другие везикулы переносятся к аппарату Гольджи и сливаются с ним. От аппарата Гольджи также отшнуровываются мембранные пузырьки: одни образуют лизосомы, другие сливаются с наружной мембраной. Внутри этих пузырьков переносятся и белки.
Пожалуй, одной из важнейших функций всего ЭПР является синтез клеточных мембран. Мембраны всех одномембранных клеточных органоидов (т. е. кроме мембран митохондрий и пластид), синтезируются в ЭПР. Ядерная мембрана, можно сказать, образована самим эндоплазматическим ретикулумом. К другим органоидам мембранный материал доставляется благодаря тому, что от ЭПР все время отшнуровываются небольшие мембранные пузырьки. Часть вакуолей имеет ретикулярное происхождение, то есть они образуются путем отшнуровывания от эндоплазматического ретикулума. Другие везикулы переносятся к аппарату Гольджи и сливаются с ним. От аппарата Гольджи также отшнуровываются мембранные пузырьки: одни образуют лизосомы, другие сливаются с наружной мембраной. Внутри этих пузырьков переносятся и белки.
Отшнуровывание мембранных пузырьков от цистерн ЭПР
и слияние их с пузырьками аппаратаГольджи (АГ)
Еще одной важной функцией эндоплазматического ретикулума является модификация белков. Расположенные здесь ферменты «умеют», например, присоединять к белкам углеводные остатки, формируя гликопротеины.
Таким образом, основными функциями ЭПР являются:
- компартментализация – разделение клетки на отсеки;
- «ферментативный конвейер» – мембрана ЭПР является поверхностью, с закрепленными на ней ферментами, на которой происходят все внутриклеточные процессы;
- биосинтез и модификация белков, липидов, углеводов;
- транспортировка различных веществ;
- депонирование различных веществ.
- формирование прочих одномембранных органоидов;
Комплекс Гольджи(аппарат Гольджи, пластинчатый комплекс).
Назван в честь Камилло Гольджи, который в 1898г. обнаружил его в нервных клетках. Он представляет собой целую систему мембранных пузырьков, анатомически не образующих внутри клетки единого пространства. Внутренность этих пузырьков нигде не соединяется с цитоплазмой, но, в отличие от эндоплазматической сети, не образует и единой непрерывной системы: связь между разными частями аппарата Гольджи осуществляется благодаря тому, что от одних его частей отшнуровываются мембранные пузырьки и сливаются с другими частями. Комплекс Гольджи обычно располагается около ядра.
Структурной единицей аппарата Гольджи является диктиосома – стопка из 3-13 уплощенных одномембранных замкнутых дискообразных цистерн диаметром 0,2-0,5 мкм. Количество самих диктиосом, формирующих в клетке аппарат Гольджи, может быть от одной до 20. Монодиктиосомный комплекс Гольджи более характерен животным клеткам, полидиктиосомный – клеткам растений и грибов, хотя подобная корреляция не является строгой.
 Диктиосомы обладают полярностью (не симметричны). Полюс формирования («наружная» поверхность) представлен множеством везикул, отшнуровавшихся от ЭПР. Выше эти везикулы сливаются, образуя цистерны. Цистерны изогнуты, по краям имеют вздутия, и своей вогнутой стороной обращены в сторону секреторного полюса («внутренняя» поверхность). В центральной части комплекса Гольджи от цистерн отшнуровываются первичные лизосомы. На секреторном полюсе из цистерн образуются секреторные везикулы, которые направляются к плазмалемме и сливаются с ней, выплескивая содержимое за пределы клетки. Таким образом, комплекс Гольджи – это структура, пребывающая в постоянной динамике: всё новые цистерны образуются на полюсе формирования, оттесняя вышележащие к секреторному полюсу, где из них формируются секреторные везикулы.
Диктиосомы обладают полярностью (не симметричны). Полюс формирования («наружная» поверхность) представлен множеством везикул, отшнуровавшихся от ЭПР. Выше эти везикулы сливаются, образуя цистерны. Цистерны изогнуты, по краям имеют вздутия, и своей вогнутой стороной обращены в сторону секреторного полюса («внутренняя» поверхность). В центральной части комплекса Гольджи от цистерн отшнуровываются первичные лизосомы. На секреторном полюсе из цистерн образуются секреторные везикулы, которые направляются к плазмалемме и сливаются с ней, выплескивая содержимое за пределы клетки. Таким образом, комплекс Гольджи – это структура, пребывающая в постоянной динамике: всё новые цистерны образуются на полюсе формирования, оттесняя вышележащие к секреторному полюсу, где из них формируются секреторные везикулы.
Электронная микрофотография и схема диктиосомы в вертикальном разрезе (увеличение фотографии – х50 000).
 На схеме: стрелка – направление развития диктиосомы, красный цвет – модифицированные мембраны.
На схеме: стрелка – направление развития диктиосомы, красный цвет – модифицированные мембраны.
Функции комплекса Гольджи:
 1) Модификация мембран. Здесь в состав мембран включаются необходимые мембранные белки и модифицируются мембранные липиды. Как вы помните, плазмалемма на 2-3нм толще мембран ЭПР, и прямой контакт между ними невозможен. Однако комплекс Гольджи, образующийся из везикул ЭПР, на секреторном полюсе образует везикулы, сливающиеся с плазмалеммой. Это возможно благодаря постепенной модификации липидного состава мембран в комплексе Гольджи (см. схемы справа).
1) Модификация мембран. Здесь в состав мембран включаются необходимые мембранные белки и модифицируются мембранные липиды. Как вы помните, плазмалемма на 2-3нм толще мембран ЭПР, и прямой контакт между ними невозможен. Однако комплекс Гольджи, образующийся из везикул ЭПР, на секреторном полюсе образует везикулы, сливающиеся с плазмалеммой. Это возможно благодаря постепенной модификации липидного состава мембран в комплексе Гольджи (см. схемы справа).
2) Рост плазмалеммы, а, следовательно, и всей клетки.
3) Модификация белков. Этот процесс похож на такую же функцию эндоплазматического ретикулума, хотя конкретные химические реакции, идущие в этих двух органоидах, различаются.
4) Формирование первичных лизосом.
5) Клеточная секреция. Две последние функции тесно связаны еще с одной, крайне важной:
6) Сортировка белков. Этот процесс происходит по двум принципам: функциональному и пространственному.
а) Функциональная сортировка. От аппарата Гольджи отшнуровываются мембранные пузырьки, из которых одни сливаются с плазмалеммой, а другие превращаются в лизосомы. Естественно, в них должны попадать разные белки, и аппарат Гольджи сортирует белки в соответствии с их функцией: лизосомальные – в лизосомы, секретируемые – в секреторные пузырьки. Как это происходит?
- Все белки, которые будут транспортироваться в лизосомы, имеют в своем составе определенную последовательность аминокислот. Эта последовательность служит как бы сигналом: «поместите меня в лизосому».
- В полостях аппарата Гольджи имеется специальный фермент, который узнает лизосомальную последовательность и прикрепляет к белку моносахаридный остаток – маннозо-6-фосфат.
- На мембране аппарата Гольджи есть особый белок – рецептор маннозо-6-фосфата. Белки с маннозо-6-фосфатом (т. е. будущие лизосомальные) связываются с рецептором. Рецепторы маннозо-6-фосфата вместе со связанными белками попадают в специальные пузырьки, которые отшнуровываются от мембраны – незрелые лизосомы.
 - На мембране лизосом имеется фермент, закачивающий в них ионы водорода Н+ (разумеется, закачивает он их за счет энергии АТФ).
- На мембране лизосом имеется фермент, закачивающий в них ионы водорода Н+ (разумеется, закачивает он их за счет энергии АТФ).
- Внутри пузырька среда становится кислой (рН около 4,5-5), а в кислой среде лизосомальные белки отделяются от рецептора маннозо-6-фосфата. Теперь лизосома уже готова к работе, но на мембране еще остаются рецепторы маннозо-6-фосфата. Было бы непростительным расточительством терять их при образовании каждой новой лизосомы, и природа такого расточительства не допустила:
- рецепторы маннозо-6-фосфата собираются в определенных участках мембраны лизосомы, от нее отшнуровываются специальные пузырьки с этими рецепторами, которые затем сливаются с мембранами аппарата Гольджи, и рецепторы готовы к новому использованию для образования следующей лизосомы.
Задание.
В клетке произошла мутация, в результате которой все лизосомальные ферменты перестали попадать в лизосомы; однако если в полость аппарата Гольджи ввести лизосомальные ферменты из здоровой клетки, то они окажутся в лизосомах.
Какой фермент был поврежден в результате мутации?
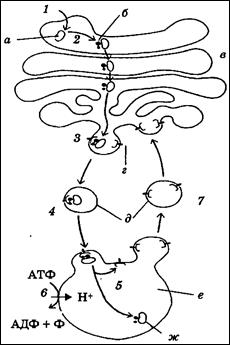
Транспорт белков в лизосомы
а – будущий лизосомный блок; б – маннозо-6-фосфат; в – аппарат Гольджи;
г – рецептор маннозо-6-фосфата; д – отшнуровавшийся от аппарата Гольджи пузырек; е – первичная лизосома; ж – зрелый лизосомный белок
1 – транспорт из ЭПР; 2 – присоединение маннозо-6-фосфата; 3– связывание с рецептором маннозо-6-фосфата; 4 – рецепторзависимый транспорт;
5 – отделение белка от рецептора при кислом рН; 6 – АТФ-зависимый транспорт ионов Н+ в лизосому; 7 – возвращение рецептора маннозо-6-фосфата
б) Пространственная сортировка. Иногда клетки имеют различающиеся полюса. Например, в клетках эпителия кишечника одна сторона обращена в просвет кишки, а другая – к кровеносному сосуду. Их поверхности сильно различаются даже внешне (одна покрыта ворсинками, другая – нет); естественно, различаются и белки, расположенные на разных поверхностях. Аппарат Гольджи как раз и сортирует белки пространственно: одни отправляет к одному полюсу, другие – к другому. Впрочем, процесс пространственной сортировки в разных клетках протекает по-разному: в некоторых клетках белки распределяются по полюсам уже на мембране, тогда как в других главная роль принадлежит аппарату Гольджи.
7) Специфические функции в отдельных клетках. У одноклеточных организмов расширенные цистерны аппарата Гольджи образуют сократительные вакуоли. В передней части сперматозоидов расширенная цистерна аппарата Гольджи образует акросому, которая содержит литические ферменты, растворяющие оболочки яйцеклетки.
Лизосомы
Лизосомы открыл биохимик Де Дюв (1955г.).
Лизосомы («лизио» – растворяю и «сома» – тело) – одномембранные органоиды, рассеянные по цитоплазме и имеющие форму пузырьков разнообразной формы и размера, диаметром до 2 мкм. Количество лизосом в клетке колеблется от 10 до 100 и более. Их главная функция – клеточное пищеварение. Лизосомы характерны для клеток животных и грибов. У растений они не выявлены – их функции выполняют вакуоли. Лизосомы, содержат около 40 различных протеолитических ферментов (протеазы, нуклеазы, липазы и кислые фосфатазы). Лизосомальные ферменты активны только в кислой среде, поддерживаемой работой протонных насосов мембраны лизосом.
Как мы уже знаем, при отшнуровывании от периферической части аппарата Гольджи образуются мелкие первичные лизосомы (около 0,1мкм). Их дальнейшая судьба может быть различна, что связано с четырьмя основными выполняемыми лизосомами функциями:
Функции собственно первичных лизосом.
1) Экзоцитоз лизосомальных ферментов. При этом ферменты выделяются в окружающую среду и расщепляют вещества за пределами клетки. Примером могут служить сапрофитное питание грибов или деятельность остеокластов – специфических клеток костной ткани, формирующих полости трубчатых костей, разрушая ткань.
2) Автолиз – лизосомы могут использоваться для саморазрушения клетки. Вы помните, что в процессе метаморфоза головастика в лягушку у него исчезает хвост. Хвост не отваливается от головастика, просто в клетках хвоста нарушается целостность ли-зосомальных мембран, ферменты лизосом выходят в цитоплазму и переваривают клетку изнутри. Автолиз является конечным этапом сложного многоэтапного процесса – апоптоза – генетически запрограммированной клеточной гибели. Апоптоз непременно сопровождает онтогенез сложноорганизованных многоклеточных животных. Так разрушаются плавательные перепонки и хвост у зародыша человека.
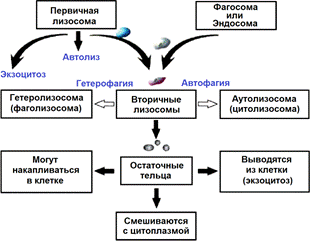
Функции, связанные с формированием вторичных лизосом (пищеварительных вакуолей).
3) Гетерофагия. Как вы уже знаете, многие животные клетки обладают способностью к фагоцитозу, в ходе которого образуется фагосома (от 250 нм и больше). К аналогичному процессу – пиноцитозу, сопровождающемуся образованием более мелких эндосом (до 150 нм), способны почти все эукариотические клетки. После образования фагосомы или эндосомы к ней подходит первичная лизосома и эти два мембранных пузырька сливаются. Образовавшийся органоид носит название гетеролизосомы (фаголизосомы) и является одной из двух разновидностей вторичных лизосом.
4) Автофагия. Лизосомы могут расщеплять не только частицы, попавшие извне, – они способны переваривать собственные «состарившиеся» и неработающие органоиды, например митохондрии, рибосомы и т.д. При этом на первом этапе органоид обволакивается мембраной ЭПР, а далее, образовавшийся везикул сливается с первичной лизосомой. Образуется вторая разновидность вторичных лизосом – аутолизосома (цитолизосома). Аутолизосомы выполняют роль внутриклеточных чистильщиков, их количество возрастает при повреждении клеток, при стрессах, при различных генетических и инфекционных заболеваниях. Каким образом лизосомы различают органоиды «плохие» и «хорошие» – неизвестно.
 Во вторичной лизосоме есть пищеварительные ферменты, кислая среда (все это благодаря первичной лизосоме) и макромолекулы, которые можно расщеплять (крупные пищевые частицы из фагосомы, растворенные во внеклеточной жидкости молекулы, попавшие в эндосому или клеточные структуры). Пищеварительные ферменты будут расщеплять те макромолекулы, которые оказались во вторичной лизосоме, а продукты расщепления – аминокислоты, моносахариды и прочие полезные вещества – будут транспортироваться через лизосомальную мембрану в цитоплазму клетки.
Во вторичной лизосоме есть пищеварительные ферменты, кислая среда (все это благодаря первичной лизосоме) и макромолекулы, которые можно расщеплять (крупные пищевые частицы из фагосомы, растворенные во внеклеточной жидкости молекулы, попавшие в эндосому или клеточные структуры). Пищеварительные ферменты будут расщеплять те макромолекулы, которые оказались во вторичной лизосоме, а продукты расщепления – аминокислоты, моносахариды и прочие полезные вещества – будут транспортироваться через лизосомальную мембрану в цитоплазму клетки.
Лизосома, содержащая непереваренные вещества, превращается в остаточное тельце. Остаточные тельца выводятся из клетки путем экзоцитоза, смешиваются с цитоплазмой в виде включений или накапливаются вплоть до гибели клетки. В последнем случае они могут вызывать различные патологические процессы (у человека известно около 12 врожденных заболеваний, при которых отмечается дисфункция лизосом).
Таким образом, различают 4 функциональных вида лизосом:
Первичная лизосома – содержит неактивные ферменты.
Два вида вторичных лизосом (пищеварительных вакуолей):
- гетеролизосома (фаголизосома) – возникает как результат соединения первичной лизосомы с фагосомой или эндосомой
- аутолизосома (цитолизосома) – образуются при соединении первичной лизосомы с компонентами самой клетки.
Остаточное тельце – содержит непереваренные вторичными лизосомами питательные вещества.
Задание.
1) Раствор аммиака сильно угнетает способность лизосом переваривать попавшие в них частицы. Почему?
2) Какую проблему вы видите в том, что лизосомы способны одновременно и к автофагии и к гетерофагии? Как эта проблема решена в клетке?
Прочие одномембранные органоиды
Секреторные вакуоли – это короткоживущие одномембранные пузырьки, которые образуются путем отшнуровывания от периферической части аппарата Гольджи. Секреторные вакуоли содержат разнообразные вещества (неактивные ферменты, или проферменты, медиаторы нервной системы, гормоны различной природы, полисахариды, липиды и т.п.), выводимые за пределы клетки путем экзоцитоза. Секреторные вакуоли хорошо видны в специализированных клетках экзокринных желез.
Пероксисомы(микротельца) – это одномембранные пузырьки диаметром 0,3-1,5 мкм, которые образуются путем отшнуровывания от цистерн гранулярного эндоплазматического ретикулума, а иногда сохраняют с ним связь весь период функционирования. Пероксисомы заполнены гранулярным матриксом и содержат разнообразные ферменты, например, каталазу, разлагающую пероксид водорода (2Н2О2 → 2Н2О + О2↑). В ряде случаев пероксисомы содержат и другие системы ферментов.
Сферосомы – это одномембранные пузырьки диаметром около 1 мкм, которые образуются путем отшнуровывания от эндоплазматической сети. Сферосомы характерны для клеток растений. Первичная сферосома (просферосома) накапливает липиды, увеличивается в размерах, затем утрачивает мембрану и превращается в масляную каплю. Кроме липидов в составе сферосом имеются ферменты, контролирующие превращения липидов – липазы.
Вакуоли – это заполненные жидкостью крупные одномембранные полости. Настоящие вакуоли имеются только у растений. Первично вакуоли образуются при слиянии мелких пузырьков, отшнуровывающихся от эндоплазматической сети. В ходе функционирования вакуолей в их состав могут включаться пузырьки, отшнуровывающиеся и от аппарата Гольджи. В молодой клетке растений содержится несколько вакуолей, которые в зрелой клетке сливаются в одну центральную вакуоль. Мембрана крупных вакуолей имеет собственное название – тонопласт. Содержимое вакуолей называется клеточным соком (концентрированный раствор органических кислот, сахаров, метаболитов, некоторых белков, неорганических солей, пигментов).
Функции вакуолей разнообразны:
- участие в формировании тургорного давления (осмотическое поступление воды);
- обеспечение окраски органов растений – цветков, плодов, почек;
- накопительное пространство для промежуточных продуктов обмена веществ растений (глюкоза, лимонная кислота);
- аккумуляция экскреторных веществ (пигменты, алкалоиды);
Для желающих: подробнее о вакуолях см. 10 класс «01 Особенности клеточной организации Растений и Грибов»
Двумембранные органоиды
К двумембранным органоидам относятся митохондрии и пластиды. В состав этих органоидов входит две мембраны: внешняя и внутренняя. Эти органоиды называются полуавтономными, поскольку они содержат собственные кольцевые ДНК, все типы РНК, рибосомы 70S и способны синтезировать некоторые белки. Кроме того, митохондрии и пластиды способны к самовоспроизведению (самостоятельно делятся) и никогда не образуются из других компонентов клетки. Вне клеток они существовать не могут.
Митохондрии
Митохондрии открыл Р. Альтман (1890–1894 гг.); он называл их «биобластами». Сам термин «митохондрия» предложил К. Бенда (1897-1898гг.), однако долгое время использовался термин «хондриосома». В 1920-ых гг. О. Варбург доказал, что с митохондриями связаны процессы клеточного дыхания.
Это двумембранные органоиды, главная функция которых – аэробное дыхание (кислородный этап диссимиляции, или окислительное фосфорилирование). Митохондрии служат главным источником АТФ для большинства животных клеток, а для растительных клеток они необходимы в ночное время: днем растения получают энергию АТФ в процессе фотосинтеза, ночью же они вынуждены добывать АТФ за счет окисления органических веществ кислородом. Кроме того, в митохондриях протекают и другие реакции обмена веществ, необходимые клетке.
Дополнительные функции митохондрий: регуляция водно-солевого режима, хранение питательных веществ, хранение части генетической информации и биосинтез некоторых белков.
Форма митохондрий зависит от таксономической принадлежности организмов, от тканевой принадлежности клеток и от физиологического состояния клеток. Крупные разветвленные митохондрии могут дробиться на множество мелких, а затем вновь сливаться. За счет этих преобразований число митохондрий в клетке может изменяться от одной до десятков тысяч. В соматических клетках млекопитающих обычно содержится 500-1000 митохондрий. Во многих клетках митохондрии соединены друг с другом и образуют несколько больших комплексов (а иногда один огромный), которые называют митохондрионами. Они часто располагаются в клетке рядом с местами интенсивного потребления энергии: в жгутике сперматозоида, около актино-миозиновых нитей мышечных клеток и т. п.
Наружная мембрана митохондрий более толстая – по толщине близка к плазмалемме. Она проницаема для не очень больших молекул (Mr ≤ 60 000Да) и свободно проницаема для ионов, поскольку имеет отверстия, образованные специальными белками – поринами. Наружная и внутренние мембраны митохондрий в некоторых местах соприкасаются.
 Пространство между внутренней и внешней мембранами – перимитохондриальное пространство – заполнено межмембранным матриксом. По химическому составу от близок к цитозолю, но имеет повышенную кислотность и содержит особые белковые комплексы, ответственные за перефосфорилирование нуклеотидов: часть образованной в митохондриях АТФ тратится на получение трифосфатных форм прочих нуклеотидов: АТФ + НМФ = АМФ + НТФ (Н – любой нуклеотид). Они будут использованы при биосинтезе ДНК и РНК.
Пространство между внутренней и внешней мембранами – перимитохондриальное пространство – заполнено межмембранным матриксом. По химическому составу от близок к цитозолю, но имеет повышенную кислотность и содержит особые белковые комплексы, ответственные за перефосфорилирование нуклеотидов: часть образованной в митохондриях АТФ тратится на получение трифосфатных форм прочих нуклеотидов: АТФ + НМФ = АМФ + НТФ (Н – любой нуклеотид). Они будут использованы при биосинтезе ДНК и РНК.
Внутренняя (активная) мембрана митохондрий более тонкая, поринов не содержит и абсолютно непроницаема для элементарной неконтролируемой диффузии ионов. Она образует кристы – гребневидные впячивания разнообразной формы, которые увеличивают поверхность внутренней мембраны.
В состав внутренней мембраны входят многочисленные комплексы АТФ-синтетаз, образующие на обращенной в матрикс поверхности грибовидные тела. Эта мембрана работает как настоящая электростанция: образовавшиеся при окислении веществ возбужденные электроны (ē) передаются белкам этой мембраны (ē-транспортная цепь), которые используют энергию ē для перекачивания протонов (Н+) в перимитохондриальное пространство. Разность концентраций ионов по две стороны мембраны создает протонный потенциал (разность зарядов). Когда он достигает величины ≈220мВ, открывается канал в АТФ-синтетазе, протоны проходят через него внутрь митохондрии (в матрикс), а энергия их движения используется ферментом для синтеза АТФ.
Детальнее эти процессы будут рассмотрены в 10 классе.
Содержимое митохондрии, отграниченное внутренней мембраной, заполнено основным веществом – матриксом. Это высококонцентрированная смесь ферментов, позволяющих окислять органические вещества (здесь реализуется цикл Кребса и цикл окисления жирных кислот). Кроме того в матриксе содержатся: митохондриальные кольцевые ДНК, все виды РНК, рибосомы 70S, ферменты биосинтеза НК и белков, а также включения. Длина кольцевой митохондриальной ДНК человека составляет 15569 пн. В ней закодировано 13 белков, 22 типа тРНК и 2 типа рРНК.
У аэробных прокариот митохондрий нет. Их функции выполняют мезосомы. Роль перимитохондриального пространства играет зазор между плазмалеммой и клеточной стенкой. Митохондрии также отсутствуют у некоторых анаэробных паразитических Простейших, но это вторичная утрата «ненужного» органоида.
Пластиды
Пластиды описал Антонио ван Левенгук (1676), но их подробное изучение связано с развитием биохимии и электронной микроскопии. Это двумембранные органоиды исключительно растений и миксотрофных протист, выполняющие разнообразные функции.
 Разновидности пластид. Пластиды различаются по форме, размерам, строению и функциям. Механизмы увеличения числа пластид в клетке разнообразны: деление, дробление, почкование. Исходной формой пластид являются бесцветные пропластиды. Увеличиваясь в размерах, пропластиды превращаются в бесцветные лейкопласты. Лейкопласты в своем развитии превращаются или в зеленые хлоропласты, или в бесцветные запасающие пластиды: амилопласты содержат крахмал; липидопласты накапливают липиды; протеинопласты накапливают белки и становятся их хранилищем. Хромопласты представляют собой последнюю стадию существования хлоропластов: в них происходит разрушение зеленых хлорофиллов, но длительное время сохраняются желтые и красные пигменты.
Разновидности пластид. Пластиды различаются по форме, размерам, строению и функциям. Механизмы увеличения числа пластид в клетке разнообразны: деление, дробление, почкование. Исходной формой пластид являются бесцветные пропластиды. Увеличиваясь в размерах, пропластиды превращаются в бесцветные лейкопласты. Лейкопласты в своем развитии превращаются или в зеленые хлоропласты, или в бесцветные запасающие пластиды: амилопласты содержат крахмал; липидопласты накапливают липиды; протеинопласты накапливают белки и становятся их хранилищем. Хромопласты представляют собой последнюю стадию существования хлоропластов: в них происходит разрушение зеленых хлорофиллов, но длительное время сохраняются желтые и красные пигменты.
Хлоропласты – это зеленые пластиды. Форма и количество хлоропластов в клетках относительно постоянны и зависят от таксономической принадлежности организмов, а также от уровня полиплоидии (у полиплоидов пластид больше). Обычно в клетке содержится несколько десятков хлоропластов.
Строение хлоропластов.
Внутреннее содержимое хлоропластов называется строма. Строма содержит пластидную кольцевую ДНК в виде множества копий, связанных с внутренней мембраной (в картофеле 22, в пшенице 900, у гороха 270 копий ДНК хлоропластов составляет 12% общей клеточной ДНК), РНК, рибосомы 70S и включения.
Внутренняя мембрана образует впячивания, которые называются тилакоиды (аналог крист митохондрий). Тилакоиды вскоре теряют связь с внутренней мембраной и превращаются в уплощенные цистерны, образуя третью мембрану хлоропластов – тилакоидную.
 Одиночные тилакоиды тилакоидной мембраны называют ламеллами, стопки тилакоидов (подобные стопкам монет) – гранами. Тилакоидная мембрана содержат фотосистемы (белково-пигментные комплексы с обязательным участием хлорофиллов), белки ē-транспортной цепи и многочисленные комплексы фермента АТФ-синтетазы.
Одиночные тилакоиды тилакоидной мембраны называют ламеллами, стопки тилакоидов (подобные стопкам монет) – гранами. Тилакоидная мембрана содержат фотосистемы (белково-пигментные комплексы с обязательным участием хлорофиллов), белки ē-транспортной цепи и многочисленные комплексы фермента АТФ-синтетазы.
Внутреннее содержимое тилакоидов называется люменом.
Таким образом, в хлоропласте имеются три пространственных отсека:
- межмембранное пространство,
 - занятое стромой пространство между внутренней мембраной и мембраной тилакоидов,
- занятое стромой пространство между внутренней мембраной и мембраной тилакоидов,
- занятое люменом тилакоидное пространство – внутри тилакоидов.
Главной функцией хлоропластов является проведение реакций фотосинтеза, в ходе которых:
1) получается АТФ, которую клетка может использовать на энергетические нужды;
2) из С02 и Н20 получаются органические вещества и побочный продукт фотолиза воды – О2.
Фотосинтез проходит в две стадии («световая» и «темновая» фазы фотосинтеза), но только одна из них (световая) напрямую зависит от света. Важно помнить, что темновые реакции нормально проходят как в темноте, так и на свету, и нуждаются в продуктах световой стадии. Выражение «темновые реакции» просто означает, что свет как таковой в них не участвует.
При световых реакциях энергия света возбуждает электроны хлорофилла в составе фотосистем. Эти возбужденные электроны (ē) передаются белкам ē-транспортной цепи тилакоидной мембраны, которые, как и в митохондриях, используют энергию ē для перекачивания протонов (Н+) через мембрану; в данном случае – в люмен (вещество тилакоидного пространства). Разность концентраций ионов по две стороны мембраны создает протонный потенциал (разность зарядов). Когда он достигает величины ≈220мВ, открывается канал в АТФ-синтетазе, протоны проходят через него в строму, а энергия их движения используется фер
|
из
5.00
|
Обсуждение в статье: Особенности различных клеточных мембран |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы