 |
Главная |


Органы растения: корень
|
из
5.00
|
Корень – это орган растения, обладающий неограниченным верхушечным ростом, имеющий радиальное строение и не несущий листьев.
Лист – боковой вегетативный орган растения.
Побег – стебель, с расположенными на нем листьями и почками. Это основная часть растения, состоящая из узлов и междоузлий, которая растет в длину за счет верхушечных и вставочных меристем (образовательная ткань).
Цветок – орган семенного размножения. Это укороченный, видоизмененный побег, развивается из почки. В цветке формируются половые клетки – гаметы и происходит опыление и оплодотворение.
плодНазначения плода: предохранение семян и способствование распространению. Стенки плода называют околоплодником.
Семя – зачаточное растение. Развивается из семизачатка.
Общая характеристика водорослей
10-10-2006 10:16 (0)

Общая характеристика
Особенности строения тела. Водоросли — сборная группа низших, преимущественно водных, растений. Характерной особенностью всех водорослей является то, что их тело не расчленено на вегетативные органы (корень, стебель, лист), а представлено талломом, или слоевищем. По этой причине их называют талломными, или слоевищными, растениями. В отличие от высших растений у них обычно отсутствуют ткани, а органы полового и бесполого размножения, как правило, одноклеточные. Общей для водорослей является также способность к фототрофному способу питания. Вместе с тем у некоторых водорослей наряду с автотрофным существует гетеротрофный тип питания (например, у эвгленовых).
Известно более 40 тыс. видов водорослей, которые объединяются в два полцарства — Багрянки и Настоящие водоросли. Последние подразделяются на несколько обособленных отделов (Зеленые, Харовые, Диатомовые, Золотистые, Бурые и др.), которые отличаются друг от друга по ряду таких важнейших признаков, как структура таллома, набор фотосинтезирующих пигментов и запасных питательных веществ, особенности размножения и циклы развития, местообитание и т. п.
Водорослям присуще исключительное морфологическое разнообразие. Они бывают одноклеточные (хламидомонада, хлорелла), колониальные (вольвокс) и многоклеточные, среди которых встречаются нитчатые (спирогира, улотрикс) и пластинчатые (уль-ва, ламинария и др.). Каждая из этих форм имеет различные размеры — от I мкм до нескольких десятков метров (некоторые бурые и красные). Например, бурая водоросль макроцистис грушеносный вырастает за день на 45 см и достигает длины 160 м.
Строение клетки. Клетки большинства водорослей существенно не отличаются от типичных клеток высших растений, однако у них есть свои особенности.
Клетки водоросли имеют клеточную оболочку, состоящую из целлюлозы и пектиновых веществ. У многих из них в состав клеточной стенки входят добавочные компоненты: известь, железо, альгиновая кислота и др.
Цитоплазма у большинства водорослей расположена тонким слоем вдоль клеточной стенки и окружает большую центральную вакуоль. В цитоплазме хорошо различимы эндоплазмати-ческий ретикулум, митохондрии, аппарат Гольджи, рибосомы, одно или несколько ядер.
В клетках водорослей из органелл особенно заметны хромато-форы (хлоропласты), которые в отличие от хлоропластов высших растений более разнообразны по форме, размерам, числу, строению, местоположению и набору пигментов. Они могут быть чашевидными, лентовидными, пластинчатыми, звездчатыми, дис-ковидными и др.
В хроматофорах сосредоточены фотосинтезирующие пигменты: хлорофиллы а, b, с, d,каротиноиды (каротины и ксантофиллы), фикобилины (фикоцианин, фикоэритрин). Кроме того, в матриксе хроматофора находятся рибосомы, ДНК, липидные гранулы и особые включения —пиреноиды. Пиреноиды присущи почти всем водорослям и небольшой группе мхов. Они являются не только местом скопления запасных питательных веществ, но и зоной их синтеза.
Веществами запаса у водорослей служат крахмал, масло, гликоген, волютин, водорастворимый полисахарид ламинарии и др.
Размножение. Водоросли размножаются бесполым и половым путем. Бесполое размножение происходит фрагментами колонии или таллома, а также спорами.
У одних водорослей каждая особь способна формировать и споры, и гаметы в зависимости от времени года и условий среды, у других функции бесполого и полового размножения выполняют разные особи — спорофиты (образуют споры) и гаметофиты (формируют гаметы). У многих водорослей в цикле развития наблюдается строгое чередование поколений — спорофита и гаме-тофита (красные, бурые, Некоторые зеленые).
Экологические группировки водорослей. Большинство водорослей исторически приспособились к определенным комплексам экологических факторов и образовали своеобразные сообщества, или экологические группировки: планктонные, бентосные, наземные, почвенные и др.
Планктоном называют совокупность преимущественно микроскопических свободно плавающих в толще воды организмов.
Среди них могут быть зеленые, диатомовые, золотистые, эвглено-вые водоросли. Для облегчения переноса водой планктонные водоросли имеют различные приспособления, уменьшающие плотность организмов (газовые вакуоли, включения липидов, насыщенность водой и студенистость) и увеличивающие их удельную поверхность (разветвленные выросты, приплюснутая или вытянутая форма тела и др.).
Бентосом называют водные организмы, живущие на дне водоемов или обрастающие различные водные предметы. Фито-бентос морей и океанов состоит преимущественно из бурых и красных водорослей, а в пресных водоемах представлены практически все отделы, кроме Бурых.
Обширную экологическую группировку составляют наземные, или воздушные, водоросли. Они образуют различно окрашенные налеты и пленки на коре деревьев, влажных камнях, заборах, крышах домов и т. д. Большинство из них относятся к отделам Зеленые и Диатомовые водоросли.
Почвенные водоросли живут в толще почвенного слоя. Это в основном желтозеленые, диатомовые и эвгленовые.
Кроме того, водоросли развиваются в горячих источниках, на поверхности снега и льда и т. п.
Водоросли (лат. Algae) — гетерогенная экологическая группа преимущественно автотрофных одноклеточных, колониальных или многоклеточных организмов, обитающих, как правило, в водной среде, в систематическом отношении представляющая собой совокупность многих отделов. Вступая в симбиоз с грибами, эти организмы в ходеэволюции образовали совершенно новые организмы — лишайники.
Изучение водорослей является важным этапом при подготовке специалистов в области марикультуры, рыбоводства и морской экологии. Наука о водорослях называется альгологией.
Отдел Зеленые водоросли. Общая характеристика, деление на классы, представители, роль в природе, практическое использование
Отдел включает одноклеточные, колониальные, ценобиальные и многоклеточные организмы зеленого цвета разной морфологической структуры таллома, кроме амебоидной, размером от 1 мкм до нескольких десятков сантиметров. Хроматофоры содержат хлорофилл, а- и Р- каротины, ксантофиллы: неоксантин, виолаксантин, зеаксантин, антераксантин и др. Клетки у большинства видов одеты целлюлозной, целлюлозно-пектиновой или пектиновой оболочкой, одно- или многоядерные. Хроматофоры могут быть различной формы, как правило, с пиреноидами. Запасное вещество — крахмал, редко масло. У подвижных зеленых водорослей есть глазок (стигма), 2 — 4 и более жгутиков, обычно одинаковой длины.
Практическое значение водорослей косвенного характера в наибольшей мере проявляется в рыбном, сельском и коммунальном хозяйствах, а также при эксплуатации водного транспорта и гидротехнических сооружений, отчасти в медицине, тогда как прямое использование их наиболее весомо в качестве пищевого продукта и сырья для ряда отраслей промышленности. Первое из этих направлений отражено при описании разных групп водорослей, об основных вехах второго направления речь пойдет здесь.
С древнейшего времени, уходящего своими корнями, вероятно, к первым поселениям человека на морском побережье, морские водоросли используют в пищу и как корм для домашних животных. По берегам морей и океанов их употребляют почти всюду, но особенно широко в Японии, где они являются настоящим национальным кушаньем. В европейских странах преобладает кормовое использование морских водорослей. У нас в СССР морские водоросли употребляет в пищу население главным образом дальневосточного побережья и Камчатки, в меньшей степени — жители берегов Мурмана и Белого моря и совсем не едят их на Черном море.
Употреблять в пищу можно, по-видимому, почти все водоросли, так как ядовитых форм среди них нет, не годятся лишь слишком грубые или невкусные. Так, например, на Сандвичевых островах из 115 имеющихся видов местное население употребляет в пищу около 60.
Размножение осуществляется путем деления клетки, фрагментации нити или колонии на части, а также при помощи зоо- или апланоспор. Половой процесс - изо-, гетеро-, оогамия, хологамия и конъюгация. Для некоторых водорослей характерно чередование поколений - споро- и гаметофита. Они обитают в пресных водоемах, некоторые в морях и влажных местах.
Классификация зеленых водорослей окончательно не разработана, однако чаще всего их группируют в 5 классов (вольвокофициевые, протококкофициевые, улотрихофициевые, сифонофициевые и конъюгатофициевые) по основным ступеням морфологической дифференцировки таллома, начиная от примитивных подвижных одноклеточных организмов и кончая более совершенными прикрепленными многоклеточными формами.
Спирогира (Spirogyra) — нитчатая водоросльсемейства Zygnemataceae.
Тело спирогиры — неветвящаяся нить, состоит из одного ряда цилиндрических клеток.
Размножение вегетативное (делением клеток) или половое (конъюгация). При конъюгации клетки двух соседних разнознаковых («+» или «-») нитей соединяются между собой боковыми выростами, образуется копуляционный канал, по которомупротопласт одной клетки перетекает в другую и сливается с содержимым последней. Клетка, в которой произошло слияние (зигота), закругляется, отделяется от нити и, одеваясь толстой оболочкой, превращается в зигоспору. Зигоспора перезимовывает и весной проходит мейоз, давая 4 споры, три из которых отмирают, а одна прорастает в молодую нить («+» или «-»). Все стадии, кроме зиготы и зигоспоры, - гаплоидны.
Спирогира одна из наиболее распространенных водорослей пресных вод всех частей света, встречается также и в солоноватых водах. Спирогира образует большие ватообразные скопления, которые плавают на поверхности воды или стелятся по дну и очень часто встречаются в тине стоячих и текучих вод, в прудах, болотах, канавах, речках, ручьях, бассейнах и т.д.
Вследствие большой величины клеток спирогиры, достигающей у некоторых ее видов до 0,01 мм, вследствие ясности их строения, эта водоросль является одной из наиболее хорошо изученных и служит классическим объектом при изучении анатомии клетки водоросли.
Род разделяется на 2 подрода: Euspirogyra - сливающиеся клетки одинаковы по размеру, копуляционный канал развит - и Sirogonium - сливающиеся клетки различны по размеру, копуляционный канал развит слабо или совсем отсутствует, поэтому клетки сливаются друг с другом непосредственно. Известно около 200 видов спирогир.
Высшие споровые
Подцарство высших растений объединяет многоклеточные растительные организмы, тело которых расчленено на органы – корень, стебель, листья. Их клетки дифференцированы на ткани, специализированы и выполняют определенные функции.
По способу размножения высшие растения разделяют на споровые и семенные. К споровым растениям относят мхи, плауны, хвощи, папоротники.
Мхи – это одна из самых древних групп высших растений. Представители этой группы наиболее просто устроены, их тело расчленено на стебель и листья. Они не имеют корней, а у наиболее простых – печеночных мхов даже отсутствует деление на стебель и листья, тело имеет вид слоевища. Прикрепляются мхи к субстрату и всасывают воду с растворенными в ней минеральными веществами с помощью ризоидов – выростов наружного слоя клеток. Это в основном многолетние растения небольших размеров: от нескольких миллиметров до десятка сантиметров (рис. 74).

Рис. 74. Мхи: 1 – маршанция; 2 – кукушкин лен;
 3 – сфагнум
3 – сфагнум
Для всех мхов характерно чередование поколений полового (гаметофита) и бесполого (спорофита), причем гаплоидный гаметофит преобладает над диплоидным спорофитом. Эта черта резко отличает их от остальных высших растений.
На листостебельном растении или слоевище в половых органах развиваются половые клетки: сперматозоиды и яйцеклетки.
Оплодотворение происходит только в присутствии воды (после дождялибо в половодье), по которой передвигаются сперматозоиды. Из образовавшейся зиготы развивается спорофит – спорогон с коробочкой на ножке, в которой образуются споры. После созревания коробочка вскрывается, и споры распространяются ветром. При попадании во влажную почву спора прорастает и дает начало новому растению.
Мхи – довольно распространенные растения. В настоящее время их насчитывается около 30 тыс. видов. Они неприхотливы, выдерживают сильные морозы и длительную жару, но растут только во влажных тенистых местах.
Тело печеночных мхов редко ветвится и обычно представлено листовидным слоевищем, с тыльной стороны которого отходят ризоиды. Селятся они на скалах, камнях, стволах деревьев.
В хвойных лесах и на болотах можно встретить мох – кукушкин лен. Его стебельки, усаженные узкими листьями, растут очень густо, образуя на почве сплошные зеленые ковры. К почве кукушкин лен прикрепляется ризоидами.
Кукушкин лен – растение раздельнополое, т. е. у одних особей развиваются мужские, а у других – женские половые клетки.
На женских растениях после оплодотворения образуются коробочки со спорами.
Очень широко распространены белые, или сфагновые, мхи.
Накапливая в своем теле большое количество воды, они способствуют заболачиванию почвы. Это связано с тем, что листья и стебель сфагнума, наряду с зелеными клетками, содержащими хлоропласты, имеют мертвые бесцветные клетки с порами.
Именно они и поглощают воду в 20 раз больше своей массы. Ризоиды у сфагнума отсутствуют. К почве он прикрепляется нижними частями стебля, которые, постепенно отмирая, превращаются в сфагновый торф. Доступ кислорода в толщу торфа ограничен, кроме того, сфагнум выделяет специальные вещества, предотвращающие размножение бактерий. Поэтому попавшие в торфяное болото различные предметы, погибшие животные, растения часто не сгнивают, а хорошо сохраняются в торфе.
В отличие от мхов остальные споровые имеют хорошо развитую корневую систему, стебли и листья. Более 400 млн лет назад они доминировали среди древесных организмов на Земле и образовывали густые леса. В настоящее время это немногочисленные группы в основном травянистых растений. В жизненном цикле преобладающим поколением является диплоидный спорофит, на котором образуются споры. Споры разносятся ветром и при благоприятных условиях прорастают, образуя небольшой заросток – гаметофит. Это зеленая пластинка величиной от 2 мм до 1 см. На заростке образуются мужские и женские гаметы – сперматозоиды и яйцеклетка. После оплодотворения из зиготы развивается новое взрослое растение – спорофит.
Плауны – очень древние растения. Ученые полагают, что они появились около 350–400 млн лет назад и образовывали густые леса из деревьев высотой до 30 м. В настоящее время их осталось очень мало, и это многолетние травянистые растения. В наших широтах наиболее известен плаун булавовидный (рис. 75). Его можно встретить в хвойных и смешанных лесах. Стелющийся по земле стебель плауна крепится к почве придаточными корнями.
Мелкие шиловидной формы листья густо покрывают стебель. Размножаются плауны вегетативно – участками побегов и корневищ.

Рис. 75. Папоротниковидные: 1 – хвощ; 2 – плаун;
3 – папоротник
Спорангии развиваются на прямостоячих побегах, собранных в виде колосков. Созревшие мелкие споры разносятся ветром и обеспечивают размножение и распространение растения.
Хвощи – небольшие многолетние травянистые растения. Они имеют хорошо развитое корневище, от которого отходят многочисленные придаточные корни.
Членистые стебли, в отличие от стеблей плаунов, растут вертикально вверх, от главного стебля отходят боковые побеги.
На стебле расположены мутовки очень мелких чешуйчатых листочков. Весной на зимующих корневищах вырастают бурые весенние побеги со спороносными колосками, которые после созревания спор отмирают. Летние побеги зеленые, ветвящиеся, фотосинтезируют и запасают питательные вещества в корневищах, которые перезимовывают, а весной образуют новые побеги (см. рис. 74).
Стебли и листья хвощей жесткие, пропитаны кремнеземом, поэтому животные их не едят. Хвощи растут в основном на полях, лугах, болотах, по берегам водоемов, реже в сосновых лесах. Хвощ полевой, трудноискоренимый сорняк полевых культур, используется как лекарственное растение. Стебли разных видов хвощей за счет наличия кремнезема используют как полировочный материал. Хвощ болотный ядовит для животных.
Папоротники, как хвощи и плауны, в каменноугольном периоде были процветающей группой растений. Сейчас их насчитывается около 10 тыс. видов, большинство которых распространено во влажных тропических лесах. Размеры современных папоротников колеблются от нескольких сантиметров (травы) до десятков метров (деревья влажных тропиков). Папоротники наших широт – травянистые растения с укороченным стеблем и перистыми листьями.
Под землей находится корневище – подземный побег. Из его почек над поверхностью развиваются длинные, сложные перистые листья – вайи.
Они обладают верхушечным ростом. От корневища отходят многочисленные придаточные корни.
Вайи тропических папоротников достигают в длину 10 м.
В наших краях наиболее распространены папоротники орляк, щитовник мужской и др. Весной, как только оттает почва, от корневища отрастает укороченный стебель с розеткой красивых листьев. Летом на нижней стороне листьев появляются бурые бугорки – сорусы, представляющие собой скопления спорангиев. В них образуются споры.
Молодые листья мужского папоротника используются человеком в пищу, как лекарственное растение. Вайи орляка используют для оформления букетов. В тропических странах некоторые виды папоротников разводят на рисовых полях для обогащения почвы азотом. Некоторые из них стали декоративными, оранжерейными и комнатными растениями, например нефролепис.
К споровым относятся растения, размножение и распространение которых осуществляется посредством спор, причем споры образуются двумя путями – бесполым и половым. Споровыми растениями являютсяводоросли, грибы, лишайники и высшие споровые растения (папоротники, плауны, хвощи, мхи, некоторые ископаемые растения).
В процессе эволюции около 400 млн. лет назад от многоклеточных зеленых водорослей возникли риниофиты – первые высшие растения, размножающиеся спорами, которые дали начало всем современным высшим споровым и семенным растениям. Это вымершая группа растений.В жизненном цикле высших споровых растений, как и у некоторых водорослей, чередуются особи бесполового и полового поколений, которые размножаются, соответственно, бесполым и половым путем. В полном жизненном цикле, который обеспечивает непрерывность жизни организмов, происходит чередование гаметофита (полового) и спорофита (бесполого поколения). На спорофите образуются органы бесполого размножения, на гаметофите – полового.
Высшие споровые растения после выхода на сушу в ходе эволюции претерпевали метаморфозы в двух направлениях. Так сформировались две крупные эволюционные группы – гаплоидная и диплоидная. К первой ветви относятся мхи, у которых лучше развит гаметофит, а спорофит занимает подчиненное положение. К диплоидной ветви принадлежат папоротники, хвощи и плауны. У них гаметофит редуцирован, и выглядит как заросток.
Размножение
Из спор, которые формируют особи бесполого поколения, вырастают особи полового поколения. Они имеют специальные мужские и женские половые органы, в которых развиваются мужские и женские половые клетки (гаметы) – подвижные сперматозоиды и неподвижные яйцеклетки. Для оплодотворения сперматозоид должен попасть во внешнюю среду и оплодотворить яйцеклетку, которая находится внутри женского полового органа. Для перемещения сперматозоида необходима вода. Из оплодотворенной яйцеклетки образуется зародыш. Он прорастает и превращается в особь бесполого поколения, которая размножается спорами.
Грибы, лишайники и водоросли размножаются подвижными зооспорами, спорангиеспорами, неподвижными апланоспорами. Для этих растений характерно половое размножение ооспорами, зигоспорами, вегетативное размножение также посредством спор. Из спор появляется такое же растение, как материнское. Тогда как у высших споровых растений такое растение, как то, что сформировало споры, появляется после сложного процесса оплодотворения.
Спора представляет собой клетку с двойной оболочкой. Споры у определенного вида растений могут быть одинаковыми по размерам или разными. Микроспоры – мелкие, макроспоры – крупные. Микроспоры обычно дают начало мужским заросткам, а макроспоры – женским.
Высшие споровые растения распространены в разных климатических условиях, но большинство из них обитает на влажных участках суши, так как для полового размножения им нужна вода. Однако некоторые виды этих растений встречаются и в пустынях
Отдел Голосеменные
Общая характеристика. Растения, относящиеся к отделу Голосеменные, так же как и цветковые, характеризуются тем, что производят семена.
К голосеменным растениям относятся ель, сосна, гинкго, лиственница, кедр, эфедра и др. Все они с давних пор широко используются человеком для его нужд.
Голосеменными эти растения называются потому, что их семена лежат открыто (голо) на поверхности чешуй шишек, а не в плодах, как у цветковых растений.
С помощью семян голосеменные размножаются и расселяются по земной поверхности. Семена голосеменных развиваются из семязачатков после того, как произойдет оплодотворение (слияние спермия с яйцеклеткой). Напомним, что семязачаток — это многоклеточное образование, в котором развивается яйцеклетка.
Оплодотворение внутри семязачатка, развитие там зародыша и формирование семени у взрослого растения — главное биологическое преимущество голосеменных растений перед споровыми в условиях жизни на суше.
В семенах (в отличие от спор) есть запас питательных веществ, они защищены семенной кожурой, и в них формируется зародыш нового растения. Как вы помните, зародыш семени состоит из зародышевого корня и зародышевого побега с зародышевыми листьями (семядолями). Семя — более надежный орган размножения и расселения, чем спора. Как только семя в благоприятных условиях прорастает, появившийся корешок легко укореняется. Семядоли проростка расправляются, и растение начинает самостоятельную жизнь.
Голосеменные — древесные растения: деревья, кустарники, редко лианы; трав среди голосеменных нет. У большинства из них имеются хорошо развитые ткани: фотосинтезирующие, проводящие, покровные, механические, запасающие и образовательные. Стебель голосеменных способен расти в толщину вследствие деления клеток камбия. Листья у большинства представителей голосеменных чешуевидные или игольчатые (хвоя).
Среди растений отдела Голосеменные много древних видов, как вымерших, так и современных. Наиболее знакомые и важные представители голосеменных растений — класс Хвойные.
Хвойные растения и их значение. Хвойные — самая многочисленная группа голосеменных растений. Их насчитывается свыше 550 видов. Они играют важную роль в природе: образуют леса, создавая тем самым условия жизни разнообразным представителям живого мира.
Хвойные растения участвуют в почвообразовании и имеют большое водорегулирующее и почвозащитное значение; широко используются их древесина, хвоя, семена, смолы, кора. Задача людей — сохранить биологическое разнообразие голосеменных растений.
Хвойные появились на Земле более 350 млн лет назад в Северном полушарии. Около 150 млн лет назад они стали господствующими в растительном покрове планеты. Хвойные — самые древние из всех ныне живущих семенных растений.
Среди хвойных есть низкорослые и стелющиеся формы (например, кедровый стланик — сосна карликовая или сосна горная, можжевельник казацкий), кустарники (можжевельник, эфедра) и растения-гиганты (секвойя вечнозеленая достигает 112 м в высоту и 11 м в диаметре). Такие крупные деревья, как ель, сосна, лиственница, пихта, кипарис, кедр, обычно живут по 200-400 лет, достигая в высоту 35-40 м.
Многообразие хвойных. Наиболее распространенные хвойные растения в нашей стране — ель, сосна, пихта, лиственница и можжевельник.
Сосна — светолюбивое высокоствольное дерево, произрастающее на любых почвах: песчаных, каменистых, болотных. Хвоинки сосны длиной 4-5 см по две или по пять сидят на очень кротких боковых побегах, расположенных на ветвях. Хвоинки живут 2-4 года, что определяет вечнозеленость сосны. У сосны сибирской (ее часто неправильно называют кедром) на укороченных побегах сидят по пять хвоинок. Это очень красивое дерево с белой, легкой и мягкой древесиной.
Ель в отличие от сосны — теневыносливое растение, произрастающее на плодородных почвах с достаточным увлажнением. Хвоинки ели — короткие, колючие, одиночно сидят на ветвях (рис. 100, 1), они живут 7-9 лет, некоторые виды, например ель шренка, - 13 лет. Ель — вечнозеленое растение. Ее древесина, как и у сосны, прочная и душистая. Она используется как строительный материал, как сырье для изготовления бумаги. Из хвои получают витамин C. Еловые леса значительно темнее сосновых боров. Они занимают в нашей стране огромные территории.
 Рис. 100. Ветки с шишками хвойных растений: 1 — ель; 2 — пихта; 3 — лиственница; 4 — можжевельник
Рис. 100. Ветки с шишками хвойных растений: 1 — ель; 2 — пихта; 3 — лиственница; 4 — можжевельник
Пихта похожа на ель, но имеет более темные, мягкие и плоские хвоинки (рис. 100, 2), гладкую кору на стволе. Нижние ветви, соприкасаясь с почвой, могут укореняться и образовывать новое, дочернее растение. Поэтому в некоторых местах пихтовые леса труднопроходимы.
Улиственницы хвоинки мягкие, собраны пучками по 20-40 штук на укороченных побегах (рис. 100, 3). Ежегодно осенью лиственница сбрасывает хвою как листопадное растение. Древесина лиственницы очень тяжелая (тонет в воде), долговечная, красивая, розоватого или светло-коричневого оттенка, но трудно обрабатываемая из-за твердости. На территории России лиственничные леса занимают самые большие территории.
Можжевельник не образует лесов, но как низкорослое деревце произрастает в сосновых борах и ельниках. Хвоя у можжевельника обыкновенно игольчатая, древесина очень прочная и твердая. Шишки у него сочно-мясистые, шаровидные, созревающие на 2-3-й год (рис. 100, 4). Растение это медленно растет и долго живет, оно широко используется в медицине, пищевой промышленности, парфюмерии и как декоративное.
Развитие семян у хвойных. Весной можно увидеть, что на верхушках молодых побегов сосны появляются маленькие (около 5 мм) зеленые женские шишки. У основания других молодых побегов среди хвои располагается скопление овальных мелких мужских шишек (рис. 101). На чешуях женских шишек ничем не защищенные, как голые, лежат семязачатки, в каждом из них образуется яйцеклетка. В мужских шишках развивается пыльца — совокупность пылинок, в которых позже образуются мужские половые клетки — спермии.
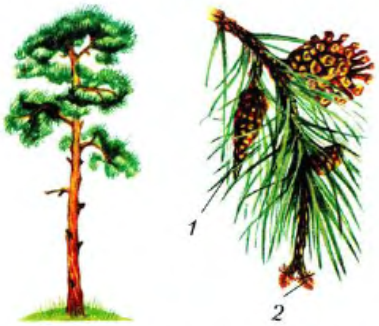 Рис. 101. Сосна обыкновенная и ветка с шишками первого и второго рода; мужские (1) и женские (2) шишкиСпермии у голосеменных — неподвижные клетки, без жгутиков. Появление такого типа мужских половых клеток — значительный этап в эволюции растений, обусловленный жизнью и развитием организмов на суше. Оплодотворение у голосеменных происходит без помощи воды. Благодаря этому голосеменные оказались способными освоить на Земле различные места обитаний, в том числе и в засушливых районах.
Рис. 101. Сосна обыкновенная и ветка с шишками первого и второго рода; мужские (1) и женские (2) шишкиСпермии у голосеменных — неподвижные клетки, без жгутиков. Появление такого типа мужских половых клеток — значительный этап в эволюции растений, обусловленный жизнью и развитием организмов на суше. Оплодотворение у голосеменных происходит без помощи воды. Благодаря этому голосеменные оказались способными освоить на Земле различные места обитаний, в том числе и в засушливых районах.
Пылинки разносятся ветром и попадают на семязачатки, голо лежащие на чешуйках женской шишки. После опыления чешуи шишки смыкаются, склеиваются смолой и начинается процесс подготовки к оплодотворению. При этом пылинка, прорастая, образует спермии и пыльцевую трубку, которая доставляет их к семязачатку, несущему яйцеклетку. Оплодотворение происходит в семязачатке лишь через 13 месяцев после опыления. У сосны шишки и семена созревают на второй год после опыления, а у ели — в тот же год.
После оплодотворения из зиготы развивается зародыш и формируются семена. Когда созреют семена, чешуи шишки раздвигаются и семена выпадают из нее. На семенах имеется пленчатое "крыло", которое позволяет им с помощью ветра далеко отлетать от материнского растения. Похожим образом размножаются все хвойные растения.
Способность голосеменных образовывать пыльцевую трубку, осуществляющую доставку неподвижных мужских гамет (спермиев) к яйцеклетке, - прогрессивный биологический шаг в эволюции растений, обусловленный жизнью в наземно-воздушной среде.
У сосны сибирской семена крупные, тяжелые, с большим запасом питательных веществ (их называют кедровыми орешками). У них нет "крыла", и распространяются они животными (кедровкой, бурундуком, белкой). Кедровые орешки — очень ценный пищевой проду
Классификация голосеменных растений
Подразделение голосеменных на классы, подклассы и порядки, а также таксономический ранг и естественность самой систематической группы в целом является предметом больших разногласий. В этой книге мы рассматриваем голосеменные как естественную группу в ранге отдела и подразделяем ее на 6 классов. Классы эти следующие:
Класс 1 — семенные папоротники, или лигиноптеридопсиды (Lyginopteridopsida, или Pteridospermae). Это древнейший класс голосеменных, геологическая история которого начинается с конца девона и заканчивается в раннем мелу; временем расцвета являются каменноугольный и пермский периоды. Листья наиболее примитивных представителей еще более или менее папоротниковидного типа, дважды или трижды перистые, с наклонностью к дихотомическому ветвлению, но в процессе эволюции они упрощаются и у своеобразного пермо-карбонового порядка глоссоптерисовых (Glossopteridales) становятся цельными. Устьица простогубые (гаплохейльные). Проводящая система представляет собой простую или рассеченную протостелу или сифоностелу. Семязачатки с пыльцевой камерой; часто бывают окружены особым плюскообразным покровом (купулой), представляющим собой видоизмененные дольки листа. Стробилы отсутствовали.
Класс 2 — саговниковые, или цикадопсиды (Cycadopsida). Ныне живущая группа голосеменных растений, геологическая история которой началась еще до позднего карбона, а временем расцвета был юрский период. Происхождение, вероятно, от семенных папоротников. Листья широкие и перистые (у бовении дважды перистые), более или менее папоротниковидные. Устьица простогубые (гаплохейльные). Стебли имеют толстую сердцевину, относительно слабо развитую и рыхлую вторичную древесину и толстую кору. Такой тип строения стебля был назван маноксильн ы м (от греч. manos — рыхлый, неплотный и xylon—древесина). Спорофиллы собраны в однополые стробилы. Семязачатки с пыльцевой камерой. Сперматозоиды с многочисленными жгутиками. Архегонии развиты.
Класс 3 — беннеттитовые, или беннеттитопсиды (Bennettitopsida). Вымершая группа, существовавшая от пермского периода до позднего мела, но наибольшую роль игравшая в юрском периоде и в раннем мелу. Происхождение, вероятно, от семенных папоротников. Листья перистые или реже цельные. Устьица сложногубые (синдетохейльные). Стебли маноксильные, как у саговниковых. Спорофиллы были собраны в стробилы, обоеполые или реже однополые. Семязачатки с пыльцевой камерой; интегумент семязачатка вытянут в длинную микропилярную трубку.
Класс 4 — гнетовые, или гнетопсиды (Gnetopsida). Современные растения. Происхождение, вероятно, от беннеттитовых. Листья цельные. Устьица простогубые (гаплохейльные) у хвойника (Ephedra) и сложногубые (синдетохейльные) у остальных двух родов. Стебли у вельвичии маноксильные, а у хвойника и гнетума пикноксильные (от греч. pyknos — плотный), т. е. кора и сердцевина относительно тонкие, а вторичная древесина относительно сильно развитая и компактная. Характерно наличие сосудов. Спорофиллы собрапы в однополые стробилы. Наличие рудиментарного семязачатка в мужских стробилах вельвичии и некоторые другие данные позволяют предполагать, что однополые стробилы гнетовых произошли из обоеполых стробилов типа беннеттитовых. Семязачатки с рудиментарной пыльцевой камерой. Мужские гаметы лишены жгутиков. Архегонии развиты (хвойник) или отсутствуют. Три порядка (Ephedrales, Welwitschiales и Gnetales), каждый из которых содержит по одному семейству.
Класс 5 — гинкговые, или гинкгопсиды (Ginkgoopsida). Класс представлен в настоящее время одним видом (Ginkgo biloba), но в мезозойскую эру, особенно перед концом юрского периода, процветал. Геологическая история восходит к позднему палеозою. Происхождение, вероятно, общее со следующим классом пинопсид от семенных папоротников. Листья от дихотомически разветвленных до двулопастных или цельных; жилкование дихотомическое. Устьица простогубые (гаплохейльные). Стебли пикноксильные. Очень редуцированные спорофиллы собраны в однополые стробилы. Семязачаток с пыльцевой камерой. Сперматозоиды с многочисленными жгутиками. В классе один порядок (Ginkgoales), который в современной флоре представлен одним монотипным семейством (Ginkgoaceae).
Класс 6 — хвойные, или пинопсиды (Pinopsida). Наиболее многочисленная группа среди современных голосеменных, геологическая история которой восходит к раннему карбону. Листья у современных форм цельные, с одной жилкой или со слабо развитым дихотомическим жилкованием, но среди вымерших форм известны вильчатые листья. Устьица простогубые (гаплохейльные). Стебли пикноксильные. Редуцированные спорофиллы собраны в однополые стробилы. Семязачаток с пыльцевой камерой (вымерший подкласс кордаитиды — Cordaitidae) или пыльцевая камера редуцирована и представлена углублением на верхушке мегаспорангия (подкласс собственно хвойные, или пиниды — Pinidae). Мужские гаметы лишены жгутиков. Делится на два подкласса — вымерший подкласс кордаитиды (Cordaitidae) и современный подкласс хвойные (Pinidae).
48 Семейство СОСНОВЫЕ (Pinaceae) Трудно назвать такое семейство даже среди цветковых растений, которое по территории, им освоенной, и по накапливаемой биомассе могло бы соперничать с сосновыми. В своем распространении это обширное семейство, насчитывающее 10 или 11 родов и не менее 250 видов, почти нацело ограничено северным полушарием. Некоторые виды сосны, ели, пихты и лиственницы забираются высоко в горы и заходят за полярный круг. Единственным видом, пересекающим экватор и заходящим в южное полушарие, является сосна Меркуза (Pinus merkusii). В семействе сосновых выделяют четыре наиболее крупных рода — пихта, лиственница, ель и сосна, насчитывающие по нескольку десятков, а то и сотню (сосна) видов. Такие роды, как кедр, псевдотсуга, кетелеерия и катайя, содержат по одному или по нескольку видов, ареалы которых крайне малы. Сосновые — вечнозеленые или, реже, листопадные деревья, иногда стелющиеся кустарники. Игловидные, чешуевидные, реже узколанцетные листья их могут быть разл
|
из
5.00
|
Обсуждение в статье: Органы растения: корень |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы