 |
Главная |


Эмбриогенез млекопитающих
|
из
5.00
|
В связи с некоторыми особенностями эмбрионального развития млекопитающих животных, относящихся к отряду приматов, а также человека их эмбриогенез рассматривается отдельно от других систематических групп млекопитающих.
Эмбриональное развитие яйцекладущих млекопитающих имеет много общего с развитием пресмыкающихся и птиц, так как яйцеклетки утконоса и ехидны очень богаты желтком и имеют сходное строение. Развитие зародышей у этих видов млекопитающих идет во время насиживания или при вынашивании яйца в сумке, т. е. во внешней среде. Дробление яйцеклетки неполное, дискоидальное, с образованием дискобластулы. Гаструляция, закладка осевых органов, развитие зародышевых оболочек осуществляется почти так же, как у пресмыкающихся.
У сумчатых млекопитающих яйцеклетки мезолецитальные, по расположению желтка их можно отнести к телолецитальным яйцеклеткам, поскольку желток локализован в области вегетативного полюса. Дробление яйцеклетки полное, неравномерное, лишь на вегетативном полюсе остается нераздробленным незначительное количество желтка (рисунок 3.1).

1 – бластомер; 2–желток; 3–вторичная оболочка яйцеклетки
Рисунок 3.1–5. Дробление яйцеклетки сумчатой куницы:
На начальных стадиях дробления бластомеры располагаются в анимально-экваториальной части над желточным шаром и полностью отделены от желтка. При последующих дроблениях бластомеры занимают экваториальное положение, а желточный шар располагается в центре. С увеличением количества бластомеров они постепенно достигают полюсов, полностью обрастая желток, который оказывается в центре бластодермического пузырька, или бластоцисты, где находится уже в жидком виде.
В связи с незначительным количеством желтка в яйцеклетке процесс его обрастания у сумчатых млекопитающих проходит быстрее, чем у пресмыкающихся, птиц и яйцекладущих млекопитающих. В однослойном бластодермическом пузырьке клетки, расположенные на анимальном полюсе, несколько мельче клеток, находящихся на вегетативном полюсе. Клеточный материал анимального полюса бластоцисты является исходным для образования зародыша. Стенка бластодермического пузырька у анимального полюса путем деляминации и иммиграции отщепляет в полость бластоцисты слой клеток будущей энтодермы, а сама превращается в наружный зародышевый листок – эктодерму. На этом завершается первая фаза гаструляции.
Далее возникает первичный узелок и первичная полоска, и происходит вторая фаза гаструляции с образованием мезодермы путем инвагинации эктодермы с последующей иммиграцией клеток в пространство между эктодермой и энтодермой. Эта фаза гаструляции, а также закладка осевых органов проходят примерно так же, как у пресмыкающихся, птиц и яйцекладущих млекопитающих.
Развитие сумчатых млекопитающих имеет следующие особенности:
1) клетки внезародышевой бластодермы, т. е. наружные стенки бластодермического пузырька, образуют трофобласт – слой клеток, который, прилегая к слизистой оболочке матки, приобретает способность к всасыванию питательных веществ – так называемого маточного молочка, выделяемого железами слизистой оболочки матки.
2) возникновение плаценты примитивного типа, которая получила название полуплаценты. Этот орган является принципиально новым в процессе эволюции и формируется он не у всех сумчатых. Полуплацента представляет собой серозную оболочку, переходящую в трофобласт, на поверхности которой клетки эктодермы образуют простые неразветвленные ворсинки. Ворсинки серозной оболочки и трофобласта проникают в отверстия маточных желез, но не достигают соединительнотканной основы слизистой оболочки матки. С образованием первичных ворсинок серозная оболочка и трофобласт сумчатых превращаются в хорион, или ворсинчатую оболочку, осуществляющую связь зародыша с организмом матери.
У плацентарных млекопитающих наблюдается вторичная утрата желтка яйцеклеткой в связи с внутриутробным развитием зародышей. Яйцеклетки у этих животных олиголецитальные, дробление полное, неравномерное. Дробление яйцеклетки происходит во время ее продвижения по яйцеводу. В это время дробящаяся яйцеклетка у некоторых животных, например у грызунов, зайцеобразных, хищных, непарнопалых, покрывается довольно толстым слоем белка, который секретируют клетки яйцевода.
Первая борозда дробления проходит меридионально. Второе деление происходит спустя два-три дня и также проходит меридионально, третье дробление идет в экваториальной плоскости. Начиная со второго дробления бластомеры начинают делиться несинхронно, и после второго дробления зародыш представлен лишь тремя бластомерами. У некоторых млекопитающих уже после первых дроблений яйцеклетки наблюдается различие в строении бластомеров: одни бластомеры более темные, а другие имеют светлую, прозрачную цитоплазму. Темные бластомеры называют зародышевыми или базофильными, их цитоплазма имеет большое количество РНК. Эти бластомеры располагаются у анимального полюса морулы. Бластомеры со светлой цитоплазмой называют внезародышевыми, они располагаются у вегетативного полюса. Эта ранняя дифференциация бластомеров показывает, что темные бластомеры являются исходным материалом для образования тела зародыша, а светлые – для образования трофобласта (рисунок 3.2).

I – стадия 2 бластомеров; II – стадия 4 бластомеров; III – морула; IV–V –образование трофобласта; VI– бластоциста и первая фаза гаструляции:
1 – темные бластомеры; 2– светлые бластомеры; 3– трофобласт;
4– эмбриобласт; 5 – эктодерма; 6– энтодерма.
Рисунок 3.2–Начальные стадии эмбриогенеза млекопитающих животных
После четвертого дробления образуется морула и наблюдается перегруппировка бластомеров: один из них, более темный, смещается от анимального полюса к центру, а три темных и четыре светлых бластомера располагаются вокруг. Потом в центр перемещается еще один темный бластомер, а остальные располагаются по периферии. Начиная с этой стадии развития дробление бластомеров делается несинхронным: светлые, внезародышевые бластомеры делятся быстрее, чем темные, зародышевые бластомеры. В результате неравномерного дробления внезародышевые бластомеры наползают на зародышевые и постепенно обрастают их. С завершением обрастания темные бластомеры, или клетки эмбриобласта, оказываются внутри, а снаружи располагаются светлые бластомеры, или клетки трофобласта. В результате жизнедеятельности клеток трофобласта между ними и клетками эмбриобласта возникает полость и образуется бластоциста (бластодермический, или зародышевый, пузырек). Эмбриобласт формирует зародышевый узелок, а трофобласт – временную оболочку, которая обеспечивает питание зародыша до возникновения иных зародышевых оболочек.
Все описанные процессы осуществляются в зародыше плацентарных млекопитающих в то время, когда он представляет собой изолированную систему и не находится в обменных взаимоотношениях с материнским организмом, так как от него он отделен остатками вторичной оболочки – ее светлой зоной. С распадом остатков вторичной оболочки яйцеклетки трофобласт вступает в тесный контакт со слизистой оболочкой матки, улучшается питание зародыша и клетки бластоцисты начинают усиленно размножаться. На наружной поверхности клеток трофобласта, там, где располагается эмбриобласт, образуются выросты – ворсинки, которые погружаются в стенку матки. Этот процесс называют имплантацией. Имплантация зародыша в стенку матки у млекопитающих различных систематических групп наступает в различные сроки после оплодотворения – от первой до десятой недели. Впоследствии трофобласт соединяется с серозной оболочкой и входит в состав детского места, или плаценты.
Гаструляция у плацентарных млекопитающих протекает так же, как у пресмыкающихся, птиц и низших млекопитающих. От поверхности зародышевого узелка, которая обращена в полость зародышевого пузырька, отщепляется путем деляминации слой плоских клеток, представляющий собой энтодерму, а вся остальная клеточная масса зародышевого узелка именуется эктодермой. Энтодерма разрастается по всей внутренней поверхности трофобласта, образуя энтодермальную стенку первичного желточного мешка. Ту часть энтодермы, которая контактирует с клетками трофобласта, называют внезародышевой энтодермой, а ту, что граничит с эктодермой – зародышевой энтодермой.
После образования двуслойного зародыша клетки эктодермы растягиваются и образуют зародышевый щиток, который непосредственно продолжается в трофобласт. Трофобласт над зародышевым щитком истончается и разрывается, вследствие чего до образования амниона зародыш млекопитающих ничем не покрыт. В течение второй фазы гаструляции образуется мезодерма так же, как у птиц и низших млекопитающих, и зародыш становится трехслойным. Закладка осевых органов принципиально ничем не отличается от таковой у птиц и низших млекопитающих.
Зародышевые оболочки у плацентарных млекопитающих возникают в основном так же, как у пресмыкающихся, птиц и низших млекопитающих, однако в деталях строения, функции и возникновения некоторых зародышевых оболочек наблюдаются существенные различия (рисунок 3.3).
Во-первых, при возникновении хориона у млекопитающих, как и у птиц, вначале образуются примитивные первичные ворсинки трофобласта, которые затем сменяются сложными вторичными ворсинками хориона. Вторичные ворсинки возникают вследствие подрастания к хориону аллантоиса, срастания их мезодермы и врастания кровеносных сосудов аллантоиса в ворсинки хориона. С врастанием кровеносных сосудов ворсинки хориона становятся крупными, ветвистыми, увеличивая площадь соприкосновения зародышевых оболочек с организмом матери.
Во-вторых, зародышевые оболочки плацентарных млекопитающих отличаются от оболочек птиц тем, что аллантоис млекопитающих не является мочевым мешком, поскольку мочевина из тела зародыша выделяется через плаценту в кровь матери.

I – начальные стадии образования амниона, аллантоиса и серозной оболочки; II – образование амниона, аллантоиса и хориона с первичными ворсинками;
1 – тело зародыша; 2– туловищные складки; 3– амниотические складки; 4–амниотическая оболочка; 5 – желточный мешок; 6– аллантоис; 7 – сросшиеся серозная оболочка и трофобласт – хорион; 8–эктодерма; 9– мезодерма; 10–желточная энтодерма; 11– энтодерма аллантоиса; 12– первичные ворсинки хориона.
Рисунок 3.3–Развитие зародышевых оболочек у плацентарных млекопитающих
Часть аллантоиса млекопитающих принимает затем участие в образовании стенки мочевого пузыря. Связь между телом зародыша и плацентой осуществляется при помощи пуповины, или пупочного канатика, который состоит из протока желточного мешка, аллантоиса и его сосудов – пупочной вены, двух пупочных артерий. Снаружи пупочный канатик покрыт амниотической оболочкой (рисунок 3.4).
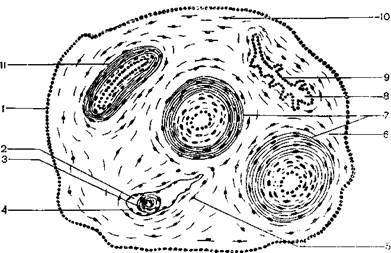
1–эктодерма амниотической оболочки; 2 – желточный мешок; 3–желточный стебелек; 4– желточная энтодерма; 5–мезодерма желточного мешка; 6–мезодерма аллантоиса; 7 – пупочные артерии; 8–аллантоис; 9–энтодерма аллантоиса; 10–мезодерма амниотической оболочки; 11–пупочная вена.
Рисунок 3.4–Пупочный канатик млекопитающего
В-третьих, у некоторых плацентарных млекопитающих, например рукокрылых, насекомоядных, приматов, амнион возникает не путем образования амниотических складок, а в результате появления полости между клетками зародышевого узелка, т. е. его образование смещается на более ранние стадии эмбрионального развития (стадия бластоцисты), еще до образования зародышевых листков. У перечисленных млекопитающих при таком способе образования амниона нет серозной оболочки, и в образовании плаценты принимает участие трофобласт, его производное – хорион, а также аллантоис.
|
из
5.00
|
Обсуждение в статье: Эмбриогенез млекопитающих |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы