 |
Главная |


Строение сердца Эндокард
|
из
5.00
|
Внутренняя оболочка сердца, эндокард (endocardium), выстилает изнутри камеры сердца, папиллярные мышцы, сухожильные нити, а также клапаны сердца. Толщина эндокарда в различных участках неодинакова. Он толще в левых камерах сердца, особенно на межжелудочковой перегородке и у устья крупных артериальных стволов - аорты и легочной артерии, а на сухожильных нитях значительно тоньше.
В эндокарде различают 4 слоя: эндотелий, субендотелиальный слой, мышечно-эластический слой и
наружный соединительнотканный слой.
Поверхность эндокарда выстлана эндотелием, лежащим на толстой базальной мембране. За ним следует субэндотелиальный слой, образованный рыхлой волокнистой соединительной тканью. Глубже располагается мышечно-эластический слой, в котором эластические волокна переплетаются с гладкими мышечными клетками. Эластические волокна гораздо лучше выражены в эндокарде предсердий, чем в желудочках. Гладкие мышечные клетки сильнее всего развиты в эндокарде у места выхода аорты. Самый глубокий слой эндокарда - наружный соединительнотканный слой - лежит на границе с миокардом. Он состоит из соединительной ткани, содержащей толстые эластические, коллагеновые и ретикулярные волокна. Эти волокна непосредственно продолжаются в волокна соединительнотканных прослоек миокарда.
Питание эндокарда осуществляется главным образом диффузно за счет крови, находящейся в камерах сердца.
Миокард
Средняя, мышечная оболочка сердца (myocardium) состоит из поперечнополосатых мышечных клеток - кардиомиоцитов. Кардиомиоциты тесно связаны между собой и образуют функциональные волокна, слои которых спиралевидно окружают камеры сердца. Между кардиомиоцитами располагаются прослойки рыхлой соединительной ткани, сосуды, нервы.
Различают кардиомиоциты трех типов:
· сократительные, или рабочие, сердечные миоциты;
· проводящие, или атипичные, сердечные миоциты, входящие в состав так называемой проводящей системы сердца;
· секреторные, или эндокринные, кардиомиоциты.
Сократительные кардиомиоцитыобразуют основную часть миокарда. Они содержат 1-2 ядра в центральной части клетки, а миофибриллы расположены по периферии. Форма клеток в желудочках - цилиндрическая, в предсердиях - неправильная, часто отросчатая.
Кардиомиоциты покрыты сарколеммой, состоящей из плазмолеммы и базальной мембраны, в которую вплетаются тонкие коллагеновые и эластические волокна, образующие "наружный скелет" кардиомиоцитов, - эндомизий. Базальная мембрана кардиомиоцитов содержит большое количество гликопротеинов. Она принимает участие в перераспределении ионов Са2+ в цикле сокращение - расслабление. Базальная мембрана латеральных сторон кардиомиоцитов инвагинирует в канальцы Т- системы (чего не наблюдается в соматических мышечных волокнах).
Кардиомиоциты желудочков значительно интенсивнее пронизаны канальцами Т-системы, чем соматические мышечные волокна. В центральной части миоцита расположено 1-2 крупных ядра овальной или удлиненной формы. Между миофибриллами располагаются многочисленные митохондрии и трубочки саркоплазматического ретикулума.
В отличие от желудочковых кардиомиоцитов предсердные миоциты чаще имеют отростчатую форму
и меньшие размеры. В миоцитах предсердий меньше митохондрий, миофибрилл, саркоплазматической сети, а также слабо развита Т-система канальцев. В тех предсердных миоцитах, где нет Т-системы, на периферии клеток, под сарколеммой, располагаются многочисленные пиноцитозные пузырьки и кавеолы. Полагают, что эти пузырьки и кавеолы являются функциональными аналогами Т-канальцев.
Между кардиомиоцитами находится интерстициальная соединительная ткань, содержащая большое количество кровеносных и лимфатических капилляров. Каждый миоцит контактирует с 2-3 капиллярами.
Секреторные кардиомиоцитывстречаются преимущественно в правом предсердии и ушках сердца. В цитоплазме этих клеток располагаются гранулы, содержащие пептидный гормон - предсердный натрийуретический фактор (ПНФ). При растяжении предсердий секрет поступает в кровь и воздействует на собирательные трубочки почки, клетки клубочковой зоны коры надпочечников, участвующие в регуляции объема внеклеточной жидкости и уровня артериального давления. ПНФ вызывает стимуляцию диуреза и натриуреза (в почках), расширение сосудов, угнетение секреции альдостерона и кортизола (в надпочечниках), снижение артериального давление. Секреция ПНФ резко усилена у больных с гипертонической болезнью.
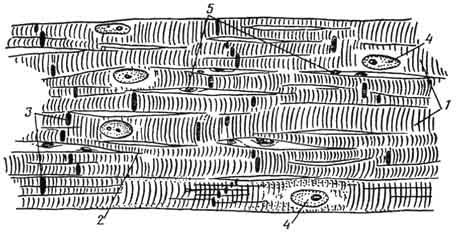
Схема строения участка рабочей мышечной ткани миокарда:
1 - миокардиоциты; 2 - анастомозы; 3 - вставочные участки; 4 - ядра миокардиоцитов; 5 - ядра
эндотелия капилляров.
Проводящие сердечные миоциты(myocyti conducens cardiacus), или атипичные кардиомиоциты, обеспечивают ритмичное координированное сокращение различных отделов сердца благодаря своей способности к генерации и быстрому проведению электрических импульсов. Совокупность атипичных кардиомиоцитов формирует так называемую проводящую систему сердца.
В состав проводящей системы входят:
· синусно-предсердный, или синусный, узел;
· предсердно-желудочковый узел;
· предсердно-желудочковый пучок (пучок Гиса) и его разветвления (волокна Пуркинье), передающие импульсы на сократительные мышечные клетки.
Различают три типа мышечных клеток, которые в разных соотношениях находятся в различных отделах этой системы.
Первый тип проводящих миоцитов - это P-клетки, или пейсмейкерные миоциты, - водители ритма. Они светлые, мелкие, отросчатые. Эти клетки встречаются синусном и предсердно-желудочковом узле и в межузловых путях. Они служат главным источником электрических импульсов, обеспечивающих ритмическое сокращение сердца. Высокое содержание свободного кальция в цитоплазме этих клеток при слабом развитии саркоплазматической сети обусловливает способность клеток синусного узла генерировать импульсы к сокращению. Поступление необходимой энергии обеспечивается преимущественно процессами анаэробного гликолиза.
Второй тип проводящих миоцитов - это переходные клетки. Они составляют основную часть проводящей системы сердца. Это тонкие, вытянутые клетки, встречаются преимущественно в узлах (их периферической части), но проникают и в прилежащие участки предсердий. Функциональное значение переходных клеток состоит в передаче возбуждения от Р-клеток к клеткам пучка Гиса и рабочему миокарду.
Третий тип проводящих миоцитов - это клетки Пуркинье, часто лежат пучками. Они светлее и шире сократительных кардиомиоцитов, содержат мало миофибрилл. Эти клетки преобладают в пучке Гиса и его ветвях. От них возбуждение передается на сократительные кардиомиоциты миокарда желудочков.
Мышечные клетки проводящей системыв стволе и разветвлениях ножек ствола проводящей системы располагаются небольшими пучками, они окружены прослойками рыхлой волокнистой соединительной ткани. Ножки пучка разветвляются под эндокардом, а также в толще миокарда желудочков. Клетки проводящей системы разветвляются в миокарде и проникают в сосочковые мышцы.
Это обусловливает натяжение сосочковыми мышцами створок клапанов (левого и правого) еще до того, как начнется сокращение миокарда желудочков.
Клетки Пуркинье- самые крупные не только в проводящей системе, но и во всем миокарде. В них много гликогена, редкая сеть миофибрилл, нет Т-трубочек. Клетки связаны между собой нексусами и десмосомами.
 Эпикард и перикард
Эпикард и перикард
Наружная, или серозная, оболочка сердца называется эпикард (epicardium). Эпикард покрыт мезотелием, под которым располагается рыхлая волокнистая соединительная ткань, содержащая сосуды и нервы. В эпикарде может находиться значительное количество жировой ткани.
Эпикард представляет собой висцеральный листок перикарда (pericardium); париетальный листок перикарда также имеет строение серозной оболочки и обращен к висцеральному слоем мезотелия. Гладкие влажные поверхности висцерального и париетального листков перикарда легко скользят друг по другу при сокращении сердца. При повреждении мезотелия (например, вследствие воспалительного процесса - перикардита) деятельность сердца может существенно нарушаться за счет образующихся соединительнотканных спаек между листками перикарда.
Эпикард и париетальный листок перикарда имеют многочисленные нервные окончания, преимущественно свободного типа.
Билет 43
1. Типы яйцеклеток и типы дробления в ряду хордовых.
В основу классификации положены 2 признака: количество и распределение желтка в я/к. По количеству желтка различают следующие виды я/к:
1. Алецитальные (безжелтковые) - у видов, развитие которых протекает с метаморфозами и эмбриональный период очень короткий или у некоторых паразитарных червей.
2. Олиголецитальный (маложелтковый) - у видов развивающихся вне организма матери в относительно благоприятной водной среде, эмбриональный период относительно короткий (пример: ланцетник). А также у видов развивающихся внутриутробно и питающихся за счет матери (пр.: млекопитающие).
3. Мезолецитальные (среднее количество желтка) - развитие вне организма матери в водной среде (пр.: лягушка).
4. Полилецитальные (многожелтковые) - развитие идет вне организма матери, причем насуше (пр.: птицы, пресмыкающиеся).
ВЫВОД: количество желтка в я/к зависит от условий где развивается зародыш, а также в какой то степени от длительности эмбрионального развития.
Вышеприведенная классификация дополняется классификацией по распределению желтка по цитоплазме:
1. Изолецитальная (равномерное распределение) - характерно для олиголецитальных я/к. Различают I изолецитальную (ланцетник) и II изолецитальную я/к (млекопитающие).
2. Телолецитальные я/к - желток распределяется по цитоплазме неравномерно, полярно - на одном полюсе (вегетативный) желток, а на другом полюсе (анимальный) ядро и органоиды. Характерно для мезо- и полилецитальных я/к; Среди телолецитальных различают 2 подгруппы:
а) умеренно телолецитальные - полярность выражено умеренно, нерезко (пр.: мезолецитальная я/к лягушки);
б) резко телолецитальные - полярность ярко выражена (пр.: птицы).
3. Центролецитальные - желток в виде узкого пояска сосредоточен вокруг ядра.
Дробление- это деление оплодотворенной я/к (уже зародыша) митозом. Дочерние клетки называются бластомерами, они не расходятся. При дроблении очень короткие интерфазы, поэтому бластомеры не успевают расти, а наоборот с каждым делением становятся размерами все меньше и меньше, т.е. количество бластомеров увеличивается, а обьем каждого отдельного бластомера уменьшается. Тип дробления зависит от типа я/к, т.е. от количества и распределения желтка.
Полное дробление- когда в дроблении участвуют все участки зародыша; характерно для олиго- изолецитальных (ланцетник, млекопитающие), а также мезо-умеренно телолецитальных я/к (лягушка).
Неполное дробление- когда дробление идет только на анимальном полюсе, вегетативный полюс перегружен желтком и в дроблении не участвует. Характерно для поли- и резко телолецитальных я/к (птицы).
Равномерное дробление- образовавшиеся бластомеры равные, одинаковые; хар-но для олиго- и I изолецитальных я/к (ланцетник).
Неравномерное дробление- образовавшиеся бластомеры неравные, разные: одни крупные, другие мелкие; одни дифференцируются в тело зародыша, другие - для питания; характерно для мезо- и полилецитальных (лягушка, птица), а также для олиго-изолецитальных я/к (млекопитающие).
Синхронное дробление- когда все бластомеры дробятся одинаковой скоростью и поэтому количество их увеличивается по правильной прогрессии, т.е. кратное увеличение; как-то: 1 : 2 : 4 : 8 и т.д.
Асинхронное дробление- кол-во бластомеров увеличивается по неправильной прогрессии; как-то: 1
: 2 : 3 : 5 - и т.д.
У ланцетника дробление полное, равномерное, синхронное. В результате такого др-ия у ланцетника образуется целобластула - полый пузырек, заполненный жидкостью. Стенка целобластулы (бластодерма) образована одним слоем бластомеров и в ней различают крышу, дно и краевую зону.
У лягушки дробление полное, неравномерное, асинхронное; в результате образуется амфибластула, состоящая из анимального и вегетативного полюса, и бластоцели с жидкостью. Бластомеры анимального полюса мелкие, дифференцируются в последующем в тело зародыша, а бл-меры вегетативного полюса крупные, перегружены желтком и обеспечивают питание зародыша.
У птиц дробление неполное (дискоидальное), неравномерное и асинхронное; в рез-те образуется дискобластула. Желток в др-ии не участвует, остается как одно целое; дроб-ие идет только на анимальном полюсе. т.е. где ядро и органоиды я/к. Образовавшиеся бл-меры распластываются на желтке и наз-ся зародышевым щитком; между зародыш. щитком и желтком имеется узкая щель - бластоцель. Зародыш.
У млекопитающих дробление полное, неравномерное, асинхронное; в рез-те образуются бл-меры 2-х типов: в центре крупные темные бл-меры - это эмбриобласт, дифф-ся в тело; по периферии мелкие светлые бл-меры - это трофобласт, участвующий при формировании хориона и плаценты. Вначале образуется морула (полости еще нет), впоследствии трофобласт всасывает жидкость слизистой яйцевыводящих путей, поэтому морула превращается в полый пузырек - эпибластула (синоним - стерробластула): стенка пузырька из одного слоя бластомеров трофобласта; полость (бластоцель) пузырька заполнена жидкостью; на одном полюсе к трофобласту изнутри прикреплен эмбриобласт. щиток
2. Мышечные ткани. Источники развития, классификации. Сердечная мышечная ткань и ее отличия от мышечной ткани соматического типа.
Классификация
I. Морфофункциональный принцип:
1. Гладкие (неисчерченные) мышечные ткани
Структурно-функциональной единицей ГМТ является гладкомышечная клетка или леомиоцит. Иннервируется вегетативной нервной системой, т.е. несознательно.
2. Поперечнополосатые (исчерченные) мышечные ткани
Структурно-функциональной единицей является мышечное волокно. Иннервируется соматической нервной системой, т.е. сознательно.
II. Гистогенетический принцип:
1. ГМТ
1) мезенхимные (из десмального зачатка в составе мезенхимы),
2) эпидермальные (из кожной эктодермы и из прехордальной пластинки),
3) нейральные (из нервной трубки),
2. ППМТ
1) Скелетные МТ - соматические (миотомные).
2) Сердечные МТ - целомические (из миоэпикардиальной пластинки висцерального листка сомита)
Поперечно-полосатая МТ соматического типа (скелетная мускулатура)-является древнейшей гистологической системой. В эмбриогенезе ПП МТ соматического типа развивается из миотомов. Структурно-функциональной единицей является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной.
Части:
1. Камбий. Миосателлитоциты - малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются.
2. Миосимпластическая часть. Мышечное волокно по форме организации живого вещества является симпластом (огромная масса цитоплазмы, где разбросаны сотни тысяч ядер).
Мышечное волокно включает большое число ядер, саркоплазму. В саркоплазме находятся: - органоиды спецназначения - миофибриллы - митохондрии - Т-система (Т-трубочки, Л-трубочки, цистерны;) - включенияя (особенно гликоген). Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Саркомер — структурная единица миофибриллы, это участок между двумя соседними телофрагмами.. Каждая миофибрилла имеет поперечные темные диски (анизотропные, полоска А, представлена толстыми миозиновыми нитями) и светлые диски (изотропные, полоски I, представлена тонкими актиновыми нитями), имеющие неодинаковое лучепреломление. Каждая миофибрилла окружена продольно расположенными и анастомозирующи- ми между собой петлями агранулярной эндоплазматической сети — саркоплазматической сети. Соседние саркомеры имеют общую пограничную структуру — Z-линию (по центру светлых И-дисков (телофрагма). По центру темных А-дисков проходит светлая зона (полоса Н), в середине которой проходит мсзофрагма (линия М). Саркомер = ½ полоски I (актин.) – полоска А (миозин.) – ½ полоски I (актин.)
Кроме сократительных белков актина и миозина в саркоплазме имеются еще вспомогательные белки - Тропонин и тропомиозин - они участвуют при обеспечении (поставке) сократительных белков ионами кальция, являющихся катализатором при взаимодействии актина и миозина. При сокращении между актиновыми и миозиновыми протофибриллами при наличии катализатора - ионов кальция образуются мостики или акто-миозиновые комплексы и это обеспечивает скольжение нитей навстречу друг к другу и укорочение саркомеров.
Канальцы саркоплазматического ретикулума располагаются в продольном направлении и образуют L-трубочки (longentidunalis = продольные); они соединяются трубочками идущими в поперечном направлении в мышечном волокне - Т-трубочками (transversus=поперечно). L- и Т-трубочки соединяются с цистернами - это своеобразные емкости для ионов кальция. В стенках цистерн имеются кальциевые насосы, откачивающие ионы Са+2 из саркоплазмы в цистерны. Нервный импульс в моторных бляшках переходит на сарколемму мышечного волокна, дальше по Т-трубочкам волна деполяризации проникает внутрь волокна, распространяется по L-трубочкам и наконец волна деполяризации проходит по стенке цистерн. В момент прохождения волны деполяризации по мембране цистерны у последней повышается проницаемость для ионов Са+2, и кальций выбрасывается в саркоплазму и подхватывается вспомогательными белками тропонином и тропомиозином и подносится к акто-миозиновому комплексу и при наличии АТФ происходит сокращение саркомера. Кальциевый насос быстро откачивает кальций обратно в цистерны - актомиозиновый комплекс распадается, поэтому происходит расслабление мышцы. Поступление нового импульса приводит к повторению всего цикла.
ПП МТ сердечного (целомического) типа- развивается из висцерального листка спланхнатомов, называемой миоэпикардиальной пластинкой.
В гистогенезе ПП МТ сердечного типа различают следующие стадии:
1. Стадия кардиомиобластов.
2. Стадия кардиопромиоцитов.
3. Стадия кардиомиоцитов.
Морфофункциональной единицей ПП МТ сердечного типа является кардиомиоцит (КМЦ). КМЦ контактируя друг с другом конец в конец формируют функциональные мышечные волокна. При этом сами КМЦ отграничены друг от друга вставочными дисками, как особыми межклеточными контактами. Морфологически КМЦ - это высокоспециализированная клетка с локализованным в центре одним ядром, миофибриллы занимают основную часть цитоплазмы, между ними большое количество митохондрий; имеется ЭПС и включения гликогена. Сарколемма (соответствует цитолемме) состоит из плазмолеммы и базальной мембраны, менее выраженной по сравнению с ПП МТ скелетного типа. В отличие от скелетной МТ сердечная МТ камбиальных элементов не имеет.В гистогенезе кардиомиобласты способны митотически делиться и в то же время синтезировать миофибриллярные белки.
Рассматривая особенности развития КМЦ, следует указать, что в раннем детстве эти клетки после разборки (т. е. исчезновения) могут вступить в цикл пролиферации с последующей сборкой акто- миозиновых структур. Это является особенностью развития сердечных мышечных клеток. Однако в последующем способность к митотическому делению у КМЦ резко падает и у взрослых практически равна нулю. Кроме того в гистогенезе с возрастом в КМЦ происходит накопление включений липофусцина. Размеры КМЦ уменьшаются.
Различают 3 разновидности КМЦ:
1. Сократительные КМЦ (типичные) - описание смотри выше.
2. Атипичные (проводящие) КМЦ - образуют проводящую систему сердца.
3. Секреторные КМЦ.
Атипичные (проводящие КМЦ - для них характерно: - слабо развит миофибриллярный аппарат; - мало митохондрий; - содержит больше саркоплазмы с большим количеством включений гликогена. Атипичные КМЦ обеспечивают автоматию сердца, так как часть их, расположенные в синусном узле сердца Р-клетки или водители ритма, способны вырабатывать ритмичные нервные импульсы, вызывающие сокращение типичных КМЦ; поэтому даже после перерезки нервов подходящих к сердцу, миокард продолжает сокращаться своим ритмом. Другая часть атипичных КМЦ проводят нервные импульсы от водителей ритма и импульсы от симпатических и парасимпатических нервных волокон к сократительным КМЦ. Секреторные КМЦ - располагаются в предсердиях; под электронным микроскопом в цитоплазме имеют ЭПС гранулярный, пластинчатый комплекс и секреторные гранулы, в которых содержится натрийуретический фактор или атриопептин – гормон, регулирующий артериальное давление, процесс мочеобразования. Кроме того секреторные КМЦ вырабатывают гликопротеины, которые соединяясь с липопротеинами крови препятствуют образованию тромбов в кровеносных сосудах.
Регенерация ПП МТ сердечного типа.Репаративная регенерация (после повреждений) - очень плохо выражена, поэтому после повреждений (пр.: инфаркт) сердечная МТ замещается соединительнотканным рубцом. Физиологическая регенерация (восполнение естественного износа) осуществляется путем внутриклеточной регенерации - т.е. КМЦ не способны делиться, но постоянно
обновляют свои изношенные органоиды, в первую очередь миофибриллы и митохондрии.
3. Спинной мозг. Строение серого и белого вещества. Собственный аппарат спинного мозга, схема его рефлекторной дуги.
Снаружи покрыт мягкой мозговой оболочкой, которая содержит кровеносные сосуды, внедряющиеся в вещество мозга.
Условно выделяют 2 половины, которые разделены передней срединной щелью и задней срединной соединительнотканной перегородкой. В центре находится центральный канал спинного мозга, который находится в сером веществе, выстлан эпендимой, содержит спинномозговую жидкость, находящуюся в постоянном движении.
По периферии располагается белое вещество, где находятся пучки нервных миелиновых волокон, которые образуют проводящие пути. Они разделены глиально-соединительнотканными перегородками. В белом веществе различают передний, боковой и задний канатики.
В средней части находится серое вещество, в котором выделяют задние, боковые (в грудных и поясничных сегментах) и передние рога. Половины серого вещества соединяются передней и задней спайкой серого вещества. В сером веществе имеются в большом количестве глиальные и нервные клетки.
Источники развития.Нервная ткань органов центральной нервной системы развивается из нервной трубки.
В сером веществе имеются в большом количестве глиальные и нервные клетки.
Нейроны серого вещества делятся на:
1) Внутренние. Полностью (с отростками) располагаются в пределах серого вещества. Являются вставочными и находятся в основном в задних и боковых рогах. Бывают:
а) Ассоциативные. Располагаются в пределах одной половины.
б) Комиссуральные. Их отростки уходят в другую половину серого вещества.
2) Пучковые нейроны. Располагаются в задних рогах и в боковых рогах. Образуют ядра или располагаются диффузно. Их аксоны заходят в белое вещество и образуют пучки нервных волокон восходящего направления. Являются вставочными.
3) Корешковые нейроны. Находятся в латеральных ядрах (ядрах боковых рогов), в передних рогах. Их аксоны выходят за пределы спинного мозга и образуют передние корешки спинного мозга.
В поверхностной части задних рогов располагается губчатый слой, где содержится большой число мелких вставочных нейронов.
Глубже данной полоски находится желатинозное вещество, содержащее в основном глиальные клетки, мелкие нейроны (последние в малом количестве).
В средней части находится собственное ядро задних рогов. Оно содержит крупные пучковые нейроны. Их аксоны идут в белое вещество противоположной половины и образуют tr. spinocerebellaris anterior и tr. spinothalamicus posterior. Клетки ядра обеспечивают экстероцептивную чувствительность.
У основания задних рогов располагается грудное ядро (столб Кларка-Штилинга), которое содержит крупные пучковые нейроны. Их аксоны идут в белое вещество этой же половины и участвуют в образовании tr. spinocerebellaris posterior и tr. spinothalamicus posterior. Клетки данного ядра обеспечивают проприоцептивную чувствительность.
В промежуточной зоне находятся латеральное и медиальное ядра. Медиальное промежуточное ядро содержит крупные пучковые нейроны. Их аксоны идут в белое вещество этой же половины и образуют tr. spinocerebellaris anterior. Обеспечивает висцеральную чувствительность.
Латеральное промежуточное ядро относится к вегетативной нервной системе. В грудном и верхнепоясничном отделах является симпатическим ядром, а в сакральном – ядром парасимпатической нервной системы. В нем содержится вставочный нейрон, который является первым нейроном эфферентного звена рефлекторной дуги. Это корешковый нейрон. Его аксоны выходят в составе передних корешков спинного мозга.
В передних рогах находятся крупные двигательные ядра, которые содержат двигательные корешковые нейроны, имеющие короткие дендриты и длинный аксон. Аксон выходит в составе передних корешков спинного мозга, а в дальнейшем идут в составе периферического смешанного нерва, представляет двигательные нервные волокна и закачивается на периферии нервно-мышечным синапсом на скелетных мышечных волокнах. Являются эффекторными. Образует третье эффекторное звено соматической рефлекторной дуги.
В передних рогах выделяют медиальную группу ядер. Она развита в грудном отделе и обеспечивает иннервацию мышц туловища. Латеральная группа ядер находится в шейном и поясничном отделах и иннервирует верхние и нижние конечности.
В сером веществе спинного мозга находится большое количество диффузных пучковых нейронов (в задних рогах). Их аксоны идут в белое вещество и сразу же делятся на две ветви, которые отходят вверх и вниз. Ветви через 2-3 сегмента спинного мозга обратно возвращаются в серое вещество и образуют синапсы на двигательных нейронах передних рогов. Данные клетки образуют собственный аппарат спинного мозга,
который обеспечивает связь между соседними 4-5 сегментами спинного мозга, за счет чего обеспечивается ответная реакция группы мышц (эволюционно выработанная защитная реакция).
Белое вещество содержит в основном миелиновые нервные волокна. Они идут пучками и образуют проводящие пути спинного мозга. Они обеспечивают связь спинного мозга с отделами головного мозга. Пучки разделяются глиальными перегородками. При этом различают восходящие пути, которые несут афферентную информацию от спинного мозга к головному. Эти пути располагаются в задних канатиках белого вещества и периферических отделах боковых канатиков. Нисходящие проводящие пути это эффекторные пути, они несут информацию от головного мозга к периферии. Располагаются в передних канатиках белого вещества и во внутренней части боковых канатиков.
|
из
5.00
|
Обсуждение в статье: Строение сердца Эндокард |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы