 |
Главная |


Современные представления о механизме белкового сплайсинга
|
из
5.00
|
Многими исследователями было показано, что белковый сплайсинг является аутокаталитическим процессом и для своего осуществления не требует присутствия ферментов или кофакторов. Однако определить точный механизм сплайсинга белков долгое время не удавалось — процесс происходит очень быстро, и обычными методами обнаружить промежуточные соединения не представлялось возможным. Главная проблема заключалась в том, что интеин в составе белка нельзя было даже выделить — сплайсинг проходил сразу после синтеза белка, и пока клетки собирались и лизировались — следов уже не оставалось. Решить эту задачу удалось группе Ф. Перлер (Francine Perler) довольно очевидным (как это теперь представляется) способом. Они изменяли ряд консервативных аминокислотных остатков в интеинах методом направленного мутагенеза. Как только мутации касались активного центра интеина — белковый сплайсинг блокировался на разных этапах, и в среде накапливались промежуточные продукты реакции. Например, изменение С-конца интеина вызывало накопление «разветвлённых» белков, у которых было... два N-конца. Исследование этих необычных белков и позволило предложить механизм белкового сплайсинга (см. рис. 3).
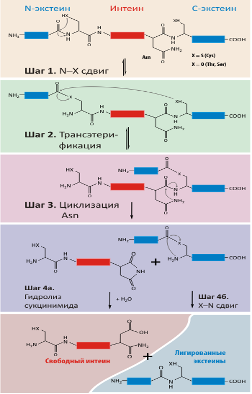
Рис. 3. Механизм белкового сплайсинга.
Событием, запускающим белковый сплайсинг, является автокаталитический N–O или N–S-сдвиг (первый аминокислотный остаток на N-конце интеина Ser или Cys, соответственно) на N-концевом сайте сплайсинга (шаг 1). В результате образуется высокореакционная эфирная или тиоэфирная связь.
С точки зрения химии, N–O/N–S-сдвиг не является энергетически выгодным процессом, поскольку в результате реакции происходит разрыв амидной (пептидной) связи и образуется высокоэнергетическая эфирная/тиоэфирная связь. Поэтому этот процесс должен катализироваться. Действительно, протекание реакции разрыва пептидной связи на N-конце облегчается как минимум двумя факторами. Во-первых, процесс катализируется самим сплайсинговым доменом интеина. Во-вторых, на эффективность N-концевого расщепления определенное влияние оказывают экстеины. Показано, что у некоторых интеинов пептидная связь, связывающая N-экстеин и первую аминокислоту интеина, находится в редкой и не характерной для белков цис-конформации. Поскольку такая связь энергетически невыгодна, ее наличие провоцирует протекание N–О или N–S сдвига, т. е. инициирует сплайсинг.
На втором этапе белкового сплайсинга происходит нуклеофильная атака образовавшейся эфирной связи OH- или SH-группой первого остатка С-экстеина. В результате происходит реакция трансэтерификации, т. е. перенос Н-концевого экстеина на боковую группу первого остатка С-экстеина (шаг 2). В результате образуется разветвленное промежуточное соединение (те самые белки с двумя N-концами, с помощью изучения которых был и предложен данный механизм сплайсинга). Такая перестановка приводит к смещению зарядов, что в свою очередь, индуцирует циклизацию боковой цепи Asn на С-конце интеина (шаг 3).
Циклизация боковой группы Asn приводит к разрыву пептидной связи между интеином и С-экстеином — разветвленная структура распадается на свободный интеин и лигированные экстеины, связанные друг с другом эфирной связью (шаг 3). Последний шаг сплайсинга белка происходит спонтанно (шаги 4а и 4б).
Согласно принятой на сегодня теории, белковый сплайсинг состоит из серии последовательных перестановок. Детальными исследованиями этих перестановок занимается российский ученый Старокадомский. Но самое удивительное, что помимо цис-сплайсинга (т. е. автокаталитического удаления интеина из белка-предшественника), у многих организмов обнаружено явление транс-сплайсинга. На пальцах это можно объяснить так: у двух белков на соответствующих концах есть по половинке интеина (назовем их интеин-подобные домены, ИПД), которые, соединяясь по типу «ключ-замок», образуют вполне функциональный интеин. А этот образованный интеин вырезает сам себя, сшивая два белка в единое целое. То есть, в результате транс-сплайсинга происходит сшивание двух белков, кодируемых двумя различными генами (рис. 4). И это не лабораторная экзотика: по такому механизму, например, происходит образование белка DnaE (одна из субъединиц ДНК-полимеразы) у Synechocystis sp.

Рис. 4. Механизм транс-сплайсинга.

Рис. 5. Предполагаемый механизм сплайсинга белков
а, б - альтернативные механизмы инициации сплайсинга, N, C - N- и C- концевые экстеины белков-предшественников
|
из
5.00
|
Обсуждение в статье: Современные представления о механизме белкового сплайсинга |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы