 |
Главная |


Как хромосома противодействует влияниям среды и избегает гибели
|
из
5.00
|
Хромосома, в основе поведения которой лежит случайность, не может избежать воздействия среды, но организованная хромосома способна к этому по той простой причине, что ей не остается ничего другого. Коль скоро хромосоме свойственна упорядоченность, ей внутренне присущи определенные правила поведения. Обладая такими собственными правилами, хромосома, очевидно, должна лишь следовать им, не поддаваясь воздействиям внешних факторов.
В основе упорядоченности этой клеточной органеллы лежит несколько механизмов.
1. Хромосома строилась в соответствии с химическим принципом самосборки. Самосборка неизбежна и иерархична; она создала собственные каналы молекулярной организации, обеспечивающие высокую степень независимости от среды.
2. Хромосома – замкнутая система. На первый взгляд эта черта хромосомы в данном контексте кажется несущественной, но она имеет решающее значение для ее эволюции. Хромосома – это не просто "нитка генов" или "кусочек ДНК"; на обоих концах ее замыкают четко выраженные особые участки – теломеры и центромеры. Обычно хромосома заканчивается на обоих концах униполярными теломерами, но у телоцентрических хромосом на одном конце эту функцию несет центромера. Без теломер хромосома гибнет. Хромосома представляет собой компартмент, а поэтому она способна сама создавать свою организацию. В хромосоме бактерий нет высокоразвитых теломер, имеющихся в хромосомах эукариот. Она разрешила проблему проще – с помощью кольцевидной формы. Кольцо – это также замкнутая система, что облегчает развитие внутренней организации.
3. Хромосома обладает целым арсеналом средств, позволяющих ей следовать собственным правилам и избегать любых посягательств на свою целостность, но в то же время она способна изменять свою структуру и функцию упорядоченным образом. Этот процесс направляется исключительно физико-химическими принципами, по которым создавалась ее первоначальная структура. К таким средствам относятся: 1) перестройки, направляющие мутационный процесс так, что допускается возникновение только данного фенотипа; 2) эффекты положения, изменяющие молекулярные функции генов; 3) наличие нуклеотидных последовательностей, способных изменять функцию и замещать другие участки ДНК, принимая на себя их функции; примером служат центромеры, способные брать на себя функцию теломер в телоцентрических хромосомах; 4) наличие транспозонов, дающих возможность хромосоме вводить участки в определенные сайты. Вместе с эписомами бактерий они позволяют вносить упорядоченность в мутации и перестройки.
Хромосомное поле выявляет строгую упорядоченность в центромеро-теломерном участке
В настоящее время все еще преобладает мнение, что в мутационном процессе и в организации хромосом главную роль играет случайность. Случайность – самая простая и самая удобная концепция, поскольку она позволяет сразу объяснить все явления, которые со строгих физико-химических позиций все еще далеко не ясны.
Недавние молекулярно-биологические исследования привлекли внимание к упорядоченным процессам построения генов и хромосомных перестроек у эукариот. Кроме того, появляются основания полагать, что мутационный процесс может направляться путем регуляции физико-химических событий, происходящих в ДНК.
Однако эукариотическая хромосома с ее гигантскими размерами все еще остается далеко не изученной на молекулярном уровне как целостная структура, т.е. как вполне определенная и строго ограниченная единица. Поэтому большинство генетиков продолжают рассматривать ее как некую случайную конструкцию.
Еще в 1950-х годах были получены данные, указывающие на жесткость структуры эукариотической хромосомы. Эта структура сформировалась в виде градиентов хромомеров, которые у более чем 70 видов начинаются у центромеры. Они сохраняют свою организацию независимо от вариаций длины хромосом. Это привело к созданию концепции хромосомного поля, согласно которой между разными участками одной хромосомы существуют определенные взаимоотношения, детерминируемые главным образом расположением центромер и теломер. В то время сведений о локализации генов было мало; не было и молекулярно-биологических методов, которые можно было бы использовать для проверки такой концепции. Однако структурные данные столь убедительно свидетельствовали в пользу жесткой и упорядоченной организации, что были сделаны следующие предсказания: 1) гены располагаются в центромеро-теломерном поле неслучайным образом; 2) каждый структурный ген и каждая последовательность ДНК стремятся занять в этом поле оптимальное место; 3) существует иерархия хромосомных участков и взаимодействие между ними, оказывающие влияние на их функцию; 4) перестройки происходят случайным образом, но следуют правилам, которые сохраняют общую структуру поля. За последние годы были собраны данные, подтвердившие справедливость этих предсказаний: 1) у более чем 700 видов, от водорослей до человека, была установлена локализация генов рРНК, которые расположены в теломерах, причем это расположение настолько регулярно, что его можно описать линейным уравнением; 2) большая часть других последовательностей ДНК, которые могут быть распознаны у большого числа видов, занимает определенное положение в пределах поля, т.е. некоторые располагаются вблизи теломер, другие вблизи центромер, а третьи – в медиальных областях плечей; 3) цитогенетические исследования на молекулярном уровне, проведенные рядом авторов, показали, что местоположение данной последовательности ДНК имеет решающее значение для определения ее функции.
Изменение положения влияет как на репликацию, так и на транскрипцию ДНК.
В одном из исследований были изучены на молекулярном уровне последовательности ДНК, участвующие в наиболее резко выраженных хромосомных перестройках среди всех, известных у высших млекопитающих. Объектом для этого исследования были выбраны олени, так как они отличаются наиболее сильной изменчивостью по числу хромосом: у самки мунтжака 2п = 6, а у самца 2п = 7; у северного оленя 2я = 70.
Последовательность ДНК оленя, состоящую из 990 пар оснований, гибридизовали по Саузерну с суммарной ядерной ДНК мунтжака. Затем из клеток благородного оленя (Cervus elaphus, 2я = 68) выделили последовательность из 770 пар оснований. Эту последовательность клонировали и гибридизовали по Саузерну с суммарной ядерной ДНК других видов Cervidae, а также гибридизовали in situ с метафазными хромосомами М. muntjak и С. elaphus. Это позволило установить, распределяется ли данная последовательность ДНК в условиях сильнейшей перестройки хромосом, произошедшей у С. elaphus, случайным образом или же она продолжает занимать свое положение в соответствии с хромосомным полем.
Оказалось, что клонированная последовательность ДНК занимает прежнее положение в участке около центромер как у С. elaphus, так и у М. muntjak. Кроме того, гены рРНК и центромеры также оставались на прежних местах, несмотря на сильную хромосомную перестройку. Результаты этих экспериментов подтверждают предсказания теории хромосомного поля, согласно которым каждая последовательность занимает оптимальное положение в центромеро-теломерном поле и стремится занять это положение после хромосомной перестройки.
Концепция хромосомного поля превратилась в теорию, потому что с течением времени она позволила сделать ряд предсказаний, подтвердившихся при экспериментальной проверке.
В свете представленных здесь данных нет оснований считать, что любая последовательность ДНК должна всегда занимать строго определенное положение в пределах поля при переносе из одной хромосомы в другую. Известная свобода перемещения необходима, иначе эволюция была бы невозможна. Теория не допускает лишь случайной перестройки последовательностей ДНК. Более того, она подчеркивает, что поведение этих последовательностей следует законам, действующим на молекулярном уровне, которые обязывают их всегда занимать мунтжака в метафазе митоза. ДНК хромосомного диска благородного оленя, содержащая 770 пар оснований, клонировали в плазмиде pBR322, осуществляли никтрансляцию в присутствии трития и гибридизовали in situ с хромосомами мунтжака. А. Клетка в метафазе; видно накопление метки в областях плечей Х-хромосомы вблизи центромеры. Б. Кариотипы трех клеток, в которых видно значительное накопление метки в участках Х-хромосомы вблизи центромеры и слабое накопление в тех же участках хромосомы I. В. Пять Х-хромосом, на которых видно, что метка накапливается не в центромере, а в участках плечей по обе стороны от нее.

Рис. 1. Семь хромосом
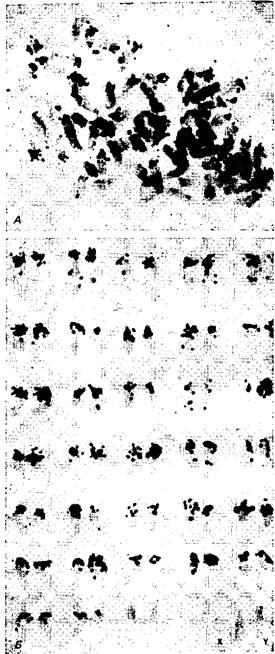
Рис. 2. 68 хромосом самца благородного оленя в метафазе митоза
Использована та же проба ДНК, меченная тритием, что и в предыдущем эксперименте– В 64 аутосомах и в хромосомах X и Y центромеры терминальные; только в двух крупных аутосомах они медиальные. А. Клетка, в которой у большинства хромосом метка накопилась вблизи центромеры. Б. Кариотип из другой клетки: 64 телоцентрические хромосомы расположены в зависимости от их длины и у всех у них метка сконцентрирована вблизи центромеры единственного плеча. В двух крупных метацентрических хромосомах в конце нижнего ряда метка отсутствует; в Х-хромосоме – самой крупной в наборе – метка также отсутствует, а в Y-хромосоме ее можно видеть вблизи центромеры.
Таблица 2. Экспериментальные данные, полученные на разных видах оленей и показывающие строгую упорядоченность хромосомных перестроек. Клонированные последовательности ДНК, центромеры и гены рРНК, несмотря на сильнейшие хромосомные перестройки, сохраняют свое местоположение в хромосомах, как и предсказывает теория хромосомного поля
| Последовательности ДНК | Расположение при большом числе хромосом | Метод | Расположение при небольшом " числе хромосом Тип распределения |
| Клонированная последовательность ДНК из 770 пар оснований Центромеры Гены рРНК | Cervus elaphus В проксимальных участках плечей вблизи центромер у 65 хромосом Muntiacus reevesi На одном конце у всех 46 хромосом Cervus elaphus. В четырех крупных хромосомах организаторы ядрышка располагаются вблизи теломер | Гибридизация по Са-узерну и гибридизация in situ Центромероспецифичная сыворотка Проба с флуоресцентной меткой Гибридизация in situ с РНК и окраска серебром | Muntiacus muntjak В проксимальных участках кие в проксимальных плечей вблизи центромер участках плечей в хромосомах X и I зи центромеры Muntiacus muntjak Вместе с другими центронируются и сохраняют мерами образуют ряды в свое местоположение виде линейно расположенных бусин, составляющих гигантские центромеры во всех хромосомах Muntiacus muntjak. Одна группа рибосом- В хромосоме I организаторных генов остается ядрышка |
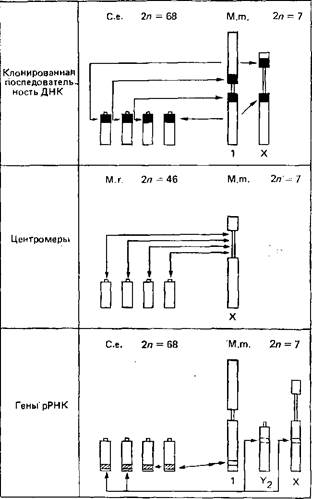
Рис. 3. Схема, в которой суммированы данные, свидетельствующие о жесткой упорядоченности хромосомных перестроек
Последовательности ДНК, центромеры и гены, кодирующие рРНК, при клонировании, несмотря на коренные хромосомные перестройки, сохраняют свое местоположение в хромосоме, что соответствует предсказаниям теории хромосомного поля. Для простоты на схеме представлены только по четыре из 68 хромосом Cervus elaphus и 46 хромосом Muntiacus reevesl. По этой же причине изображены лишь некоторые из хромосом М. muntjak. Объяснения см. табл. 2. Стрелки показывают, что перестройки могут происходить в обоих направлениях, т.е. как от высокого числа хромосом к низкому, так и в обратную сторону.
Полученные за последнее время молекулярные данные о строении генов у высших организмов не подтверждают представления о хаотичности хромосом, а указывают на строгую упорядоченность в строении генов.
У эукариот большинство генов состоит из нетранскрибируемых участков ДНК, разделенных участками, которые транскрибируются и транслируются. Число нитронов может достигать 16, как, например, у куриного гена, кодирующего белок овотрансферрин мРНК образуется путем сплайсинга РНК из экзонов. Процесс соединения участков РНК, происходящих из обособленных экзонов, отличается выраженной упорядоченностью. При сборке они соединяются соответствующими концами и в нужной последовательности.
Прерывистый ген представляет собой частный случай хромосомного поля. Первый выявляет упорядоченность в пределах гена, а второе – в пределах хромосомы.
Хромосома поддерживает постоянство, вводит новшества и производит разведку, пользуясь собственными средствами.
Хромосоме не нужен отбор для того, чтобы поддерживать постоянство, вводить новшества или производить разведку. Она содержит в себе все механизмы, необходимые для осуществления этих трех процессов. Поддержание постоянства и введение новшеств – функции антагонистические, но в мире молекул можно найти множество примеров антагонизма. Один из них – антагонизм или ингибирование гормонов, как в случае взаимно противоположных и ингибирующих действий андрогенов и эстрогенов.
Поддержание постоянства достигается путем сохранения генных последовательностей. В этом участвует несколько механизмов: 1) репарация ДНК, состоящая в замене тех оснований, которые неправильно включились или модифицировались; 2) коррекция, осуществляемая ДНК-полимеразой, которая вырезает участки ДНК, непригодные для репликации; 3) элиминация целых хромосомных участков, целых хромосом и целых хромосомных наборов. Такая элиминация – упорядоченный процесс, которому предшествуют маркировка и узнавание на молекулярном уровне.
Введение новшеств, или создание новых генных последовательностей, хорошо установлено на молекулярном уровне. Ген иммуноглобулина создан хромосомой с использованием тривиальных молекулярных механизмов. Две последовательности ДНК, которые в клетках зародышевой линии мышей непосредственно не функционировали, т.е. не транскрибировали РНК, а поэтому не могли рассматриваться как структурные гены, объединяются при помощи перестроек в соматических тканях, в результате чего они становятся активными и образуют ген иммуноглобулина. Избыточность и амплификация также представляют собой процессы, ведущие к новшествам. Они не только увеличивают число копий генов, но и порождают новые взаимодействия между существующими генами, модифицируя их активность.
Разведка заключается в переводе генетических путей на новые функциональные направления. В сущности хромосомы постоянно исследуют возможности новых решений. ДНК можно расщеплять и воссоединять. Это замечательное свойство лежит в основе всех хромосомных перестроек, выражающихся в таких странных формах, как транслокации, инверсии, дупликации и делении. Они не кажутся нам странными лишь потому, что происходят так часто в отдельных хромосомах и между ними. Новые функции, которые приобретают гены в результате некоторых из этих перестроек, приводят к образованию новых функциональных направлений. Важно отметить, что хромосома осуществляет все это в пределах собственных границ и с помощью собственных молекулярных механизмов. Для этого ей необходимо лишь оставаться в клетке, из которой она черпает необходимые молекулы.
|
из
5.00
|
Обсуждение в статье: Как хромосома противодействует влияниям среды и избегает гибели |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы