 |
Главная |


Липолиз триглицеридов в белой и бурой жировой ткани
|
из
5.00
|
ОСОБЕННОСТИ МЕТАБОЛИЗМА БЕЛОЙ ЖИРОВОЙ ТКАНИ
Энергетический обмен низкий, преимущественно анаэробный, ткань потребляет мало кислорода. Энергия АТФ в основном тратится на транспорт жирных кислот через клеточные мембраны (с участием карнитина).
Белковый обмен низкий, белки синтезируются адипоцитами преимущественно для соб-ственных нужд. На экспорт в жировой ткани синтезируются лептин, белки острой фазы вос-паления (α1-кислый гликопротеин, гаптоглобин), компоненты системы комплимента (адипсин, комплемент С3, фактор В), интерлейкины.
Углеводный обмен. Невысокий, преобладает катаболизм. Углеводный обмен в жировой ткани тесно связан с липидным.
Липидный обмен
Жировая ткань стоит на 2 месте по обмену липидов после печени. Здесь происходят реак-ции липолиза и липогенеза.
Липогенез. В жировой ткани синтез липидов идет в абсорбтивный период по глицеро-фосфатному пути. Процесс стимулируется инсулином.
Этапы липогенеза:
1. Под действием инсулина на рибосомах стимулируется синтез ЛПЛ.
2. ЛПЛ выходит из адипоцита и фиксируется на поверхности стенки капилляра с помощью гепарансульфата.
3. ЛПЛ гидролизует ТГ в составе липопротеинов
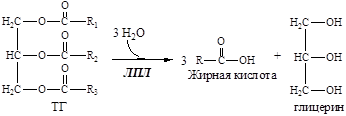
4. Образовавшийся глицерин уноситься кровью в печень.
5. Жирные кислоты из крови транспортируются в адипоцит.
6. Кроме поступающих из вне экзогенных жирных кислот, в адипоците синтезируются жир-ные кислоты из глюкозы. Процесс стимулируется инсулином.
7. Жирные кислоты в адипоците под действием Ацил-КоА синтетазы превращаются в Ацил-КоА.
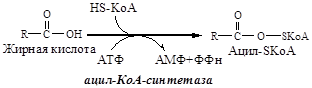
7. Глюкоза поступает в адипоцит с участием ГЛЮТ-4 (активатор инсулин).
8. В адипоците глюкоза вступает в гликолиз с образованием ФДА (активатор инсулин).
9. В цитоплазме ФДА восстанавливается глицерол-ф ДГ до глицерофосфата:
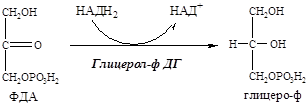
Так как в жировой ткани нет глицерокиназы, глицерофосфат образуется только из глюкозы (не может из глицерина).
10. В митохондриях глицерофосфат под действием глицеролфосфат ацилтрансферазы пре-вращается в лизофосфатид:
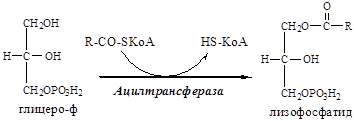
11. В митохондриях лизофосфатид под действием лизофосфатид ацилтрансферазы превраща-ется в фосфатид:
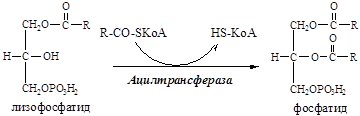
11. Фосфатид под действием фосфотидат фосфогидролазы превращается в 1,2-ДГ:
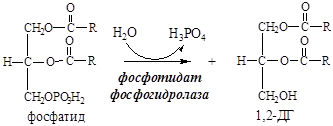
12. 1,2-ДГ под действием ацилтрансферазы превращается в ТГ:
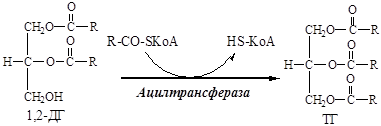
13. Молекулы ТГ объединяются в крупные жировые капли.
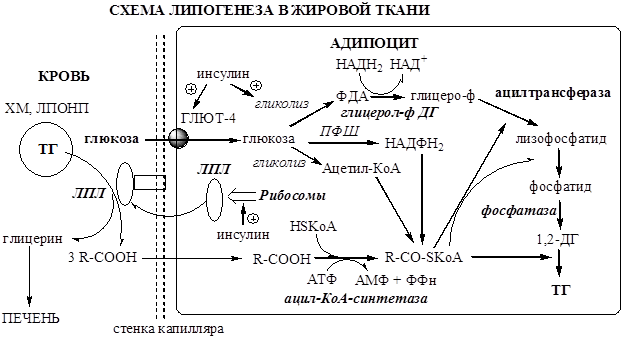
2. Липолиз. Липолиз в жировой ткани активируется при дефиците глюкозы в крови (постаб-сорбционный период, голодание, физическая нагрузка). Процесс стимулируется глюкагоном, адреналином, в меньшей степени СТГ и глюкокортикоидами.
В результате липолиза концентрация свободных жирных кислот в крови возрастает в 2 раза.
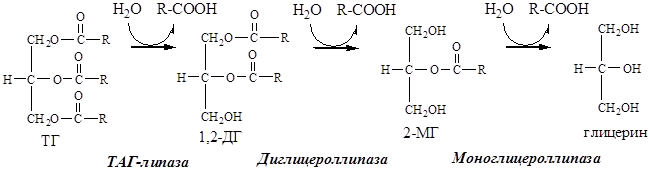

ОСОБЕННОСТИ МЕТАБОЛИЗМА БУРОЙ ЖИРОВОЙ ТКАНИ
Энергетический обмен. Ткань потребляет много кислорода, активно окисляет глюкозу и жирные кислоты. Энергетический обмен высокий. При этом, АТФ образуется только в реак-циях субстратного фосфорилирования (2 реакции гликолиза, 1 реакции ЦТК). Причина - раз-общение в митохондриях белком термогенином (РБ-1) процессов окисления и фосфорилиро-вания, низкая активность АТФ синтетазы, отсутствие дыхательного контроля со стороны АДФ. В бурой жировой ткани вся образующаяся при окислении энергия рассеивается в виде тепла (термогенез).
Термогенез в бурой жировая ткань активируется при переохлаждении СНС, а также при излишке липидов в крови, под действием лептина. Благодаря этому повышается температура тела и снижается концентрация липидов в крови. Отсутствие бурой жировой ткани у взрос-лых людей является причиной 10% всех случаев ожирения.
37. Механизмы β - окисления жирных кислот. Регуляция
β-окисление ЖК
β-окисление — специфический путь катаболизма ЖК с неразветвленной средней и корот-кой углеводородной цепью. β-окисление протекает в матриксе митохондрий, при котором от С конца ЖК последовательно отделяется по 2 атома С в виде Ацетил-КоА. β-окисление ЖК происходит только в аэробных условиях и является источником большого количества энер-гии.
β-окисление ЖК активно протекает в красных скелетных мышцах, сердечной мышце, почках и печени. ЖК не служат источником энергии для нервных тканей, так как ЖК не про-ходят через гематоэнцефалический барьер, как и другие гидрофобные вещества.
β-окисление ЖК увеличивается в постабсорбтивный период, при голодании и физической работе. При этом концентрация ЖК в крови увеличивается в результате мобилизации ЖК из жировых ткани.
Активация ЖК
Активация ЖК происходит в результате образования макроэргической связи между ЖК и HSКоА с образованием Ацил-КоА. Реакцию катализирует фермент Ацил-КоА синтетаза:
RCOOH + HSKoA + АТФ → RCO~SКоА + АМФ+ PPн
Пирофосфат гидролизуется ферментом пирофосфатазой: Н4Р2О7 + Н2О → 2Н3РО4
Ацил-КоА синтетазы находятся как в цитозоле (на внешней мембране митохондрий), так и в матриксе митохондрий. Эти ферменты отличаются по специфичности к ЖК с различной длиной углеводородной цепи.
Транспорт ЖК
Транспорт ЖК в матрикс митохондрий зависит от длины углеродной цепи.
ЖК с короткой и средней длиной цепи (от 4 до 12 атомов С) могут проникать в матрикс митохондрий путём диффузии. Активация этих ЖК происходит ацил-КоА синтетазами в матриксе митохондрий.
ЖК с длинной цепью, сначала активируются в цитозоле (ацил-КоА синтетазами на внешней мембране митохондрий), а затем переносятся в матрикс митохондрий специальной транспортной системой с помощью карнитина. Карнитин поступает с пищей или синтезируется из лизина и метионина с участием витамина С.
· В наружной мембране митохондрий фермент карнитинацилтрансфераза I (карнитин-пальмитоилтрансфераза I) катализирует перенос ацила с КоА на карнитин с образованием ацилкарнитина;
· Ацилкарнитин проходит через межмембранное пространство к наружной стороне внутренней мембраны и транспортируется с помощью карнитинацилкарнитинтранслоказы на внутреннюю поверхность внутренней мембраны митохондрий;
· Фермент карнитинацилтрансфераза II катализирует перенос ацила с карнитина на внутримитохондриальный HSКоА с образованием Ацил-КоА;
· Свободный карнитин возвращается на цитозольную сторону внутренней мембраны митохондрий той же транслоказой.
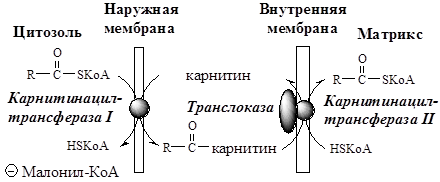
Реакции β-окисление ЖК
1. β-окисление начинается с дегидрирования ацил-КоА ФАД-зависимой Ацил-КоА дегидрогеназой с образованием двойной связи (транс) между α- и β-атомами С в Еноил-КоА. Восстановленный ФАДН2 окисляясь в ЦПЭ, обеспечивает синтез 2 молекул АТФ;
2. Еноил-КоА гидратаза присоединяет воду к двойной связи Еноил-КоА с образованием β-оксиацил-КоА;
3. β-оксиацил-КоА окисляется НАД зависимой дегидрогеназой до β-кетоацил-КоА. Восстановленный НАДН2, окисляясь в ЦПЭ, обеспечивает синтез 3 молекул АТФ;
4. Тиолаза с участием HКоА отщепляет от β-кетоацил-КоА Ацетил-КоА. В результате 4 реакций образуется Ацил-КоА, который короче предыдущего Ацил-КоА на 2 углерода. Образованный Ацетил-КоА окисляясь в ЦТК, обеспечивает синтез в ЦПЭ 12 молекул АТФ.
Затем Ацил-КоА снова вступает в реакции β-окисления. Циклы продолжаются до тех пор, пока Ацил-КоА не превратится в Ацетил-КоА с 2 атома С (если ЖК имела четное количество атомов С) или Бутирил-КоА с 3 атомами С (если ЖК имела нечетное количество атомов С).
|
из
5.00
|
Обсуждение в статье: Липолиз триглицеридов в белой и бурой жировой ткани |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы