 |
Главная |


VI. Лигазы (синтетазы)
|
из
5.00
|
КЛАССИФИКАЦИЯ И НОМЕНКЛАТУРА ФЕРМЕНТОВ
Современная классификация ферментов разработана Международным биохимическим союзом (1961г.). В ее основу положен тип катализируемой реакции и механизм. Все ферменты делят на шесть классов: I. Оксидоредуктазы. II. Трансферазы. III. Гидролазы. IV. Лиазы. V. Изомеразы. VI. Лигазы.
Каждый фермент имеет кодовый номер по классификации ферментов: первая цифра характеризует класс реакции, вторая − подкласс, третья − подподкласс, четвертая цифра указывает порядковый номер фермента в его подподклассе. Название фермента состоит из двух частей: в первой части − название субстрата (или субстратов), во второй − тип катализируемой реакции.
I. Оксидоредуктазы
Оксидоредуктазы катализируют окислительно-восстановительные реакции и подразделяются на 5 основных подклассов:
· оксидазы;
· аэробные дегидрогеназы;
· анаэробные дегидрогеназы;
· гидропероксидазы;
· оксигеназы.
Оксидазы.Катализируют удаление водорода из субстрата, используя при этом в качестве акцептора водорода только кислород. Оксидазы содержат медь, продуктом реакции является вода (исключение составляют реакции, катализируемые уриказой и моноаминоксидазой, в результате которых образуется Н2О2). Общая схема процесса следующая:
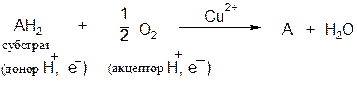
В качестве субстратов оксидаз могут выступать фенолы, полифенолы, амины. Примерами ферментов класса оксидаз являются фенолаза, цитохром-оксидаза, моноаминоксидаза, уриказа.
Фенолаза (катехолоксидаза, тирозиназа) − медьсодержащий фермент с широкой специфичностью. Он катализирует превращение монофенола
(в присутствии о-дифенола) в орто-хинон. Медь содержится также в уриказе, катализирующей окисление мочевой кислоты в аллантоин, и в моноаминоксидазе, окисляющей адреналин и тирамин в митохондриях.
1.9.3.1 Цитохромоксидаза − гемопротеин, который служит конечным компонентом дыхательной цепи и осуществляет перенос электронов на конечный акцептор − кислород. Данный фермент ингибируется оксидом углерода (II), цианидами и сероводородом. Фермент содержит две молекулы гема, в каждой из которых атом железа может переходить из состояния Fe2+ в состояние Fe3+ и обратно в ходе окисления и восстановления, а также два атома Сu, каждый из которых взаимодействует с одним из гемов.
Аэробные дегидрогеназы.Катализируют удаление водорода из суб-страта, используя в качестве акцептора водорода не только кислород, но и ис-кусственные акцепторы (например, метиленовый синий). Эти дегидрогеназы относятся к флавопротеинам. В результате реакции образуется пероксид во-дорода, а не вода:

В качестве кофермента дегидрогеназы содержат ФАД (флавинаденин-динуклеотид) или ФМН (флавинмононуклеотид). Многие флавопротеиновые ферменты содержат несколько ионов металлов.
К ферментам группы аэробных дегидрогеназ относятся: дегидрогеназа L-аминокислот (оксидаза L-аминокислот), катализирующая окислительное дезаминирование природных L-аминокислот, и ксантиндегидрогеназа (ксантиноксидаза), катализирующая окисление ксантина в мочевую кислоту. Молибденсодержащий фермент ксантиноксидаза играет важную роль в катаболизме пуриновых оснований.
Анаэробные дегидрогеназы. Катализируют удаление водорода из суб-страта, но не способны использовать кислород в качестве акцептора. Общая схема процесса следующая:
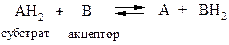
Анаэробные дегидрогеназы подразделяются в зависимости от природы кофермента на несколько групп.
НАД+-зависимые (пиридинзависимые, первичные) дегидрогеназы, содержащие в качестве кофермента НАД+ или НАДФ+. В общем случае НАД+-зависимые дегидрогеназы катализируют окислительно-восстановительные реакции специфических (например, гликолиза) и общих путей катаболизма (например, цикла Кребса, дыхательной цепи). Общая схема процесса следующая:
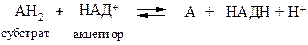
Примером НАД+-зависимых дегидрогеназ могут служить лактатдегидрогеназа, алкогольдегидрогеназа, глутаматдегидрогеназа и др. Например, 1.1.1.27 L-лактат: НАД+-оксидоредуктаза (лактатдегидрогеназа) катализирует взаимопревращение лактата и пирувата:
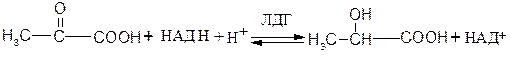
Пируват L - Лактат
ФАД- и ФМН- зависимые анаэробные дегидрогеназы (флавин-зависимые дегидрогеназы). Первичные флавинзависимые дегидрогеназы переносят восстановительные эквиваленты от субстрата непосредственно на дыхательную цепь. К ним относятся, например, 1.3.99.1 сукцинатдегидрогеназа (СДГ), ацил-КоА-дегидрогеназа и митохондриальная глицерол-3-фосфат-дегидрогеназа:
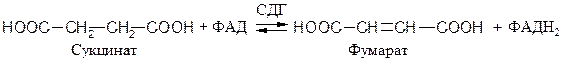
Вторичная ФМН-зависимая НАДН-дегидрогеназа − компонент дыхательной цепи, переносящий электроны от НАДН к более электроположительным компонентам:

Еще одна функция флавинзависимых дегидрогеназ − катализируемое дигидролипоилдегидрогеназой дегидрирование восстановленного липоата
(промежуточного продукта при окислительном декарбоксилировании пирувата и a-кетоглутарата). При этом вследствие низкого значения окислительно-восстановительного потенциала системы липоата переносчиком водорода от восстановленного липоата к НАД+ является флавопротеин (ФАД).
Цитохромы за исключением цитохромоксидазы классифицируются как анаэробные дегидрогеназы. В дыхательной цепи они служат переносчиками электронов от флавопротеинов к цитохромоксидазе. Цитохромы являются гемопротеинами, у которых атом железа переходит из состояния Fe2+ в Fe3+ и обратно в процессе окисления и восстановления. Помимо дыхательной цепи цитохромы имеются в эндоплазматическом ретикулуме (цитохромы Р-450 и b-5).
Гидропероксидазы.К их числу относятся ферменты пероксидаза и каталаза.
1.11.1.7 Пероксидаза − фермент, катализирующий окисление пероксидом водорода различных органических соединений − фенолов, аминов, аскорбиновой кислоты, цитохрома С. Пероксидаза обнаружена в растениях, молоке, лейкоцитах, тромбоцитах, а также в тканях, в которых происходит метаболизм эйкозаноидов. Фермент содержит протогем, который в отличие от гемовых групп большинства гемопротеинов слабо связан с апоферментом. Реакция, катализируемая пероксидазой, имеет сложный характер; суммарная реакция выглядит следующим образом:
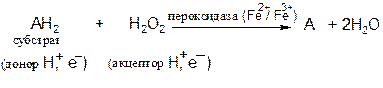
В эритроцитах глутатионпероксидаза, содержащая в качестве простетической группы селен, катализирует разложение Н2О2 и гидропероксидов липидов восстановленным глутатионом и, таким образом, защищает липиды мембран и гемоглобин от окисления пероксидами.
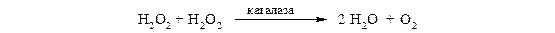 |
1.11.1.6 Каталаза − это гемопротеин, содержащий 4 гемовые группы. Наряду с пероксидазной активностью каталаза способна использовать одну молекулу Н2О2 в качестве донора электронов, а другую − в качестве акцептора электронов. В организме каталаза в основном разлагает пероксид водорода, образующийся при действии аэробных дегидрогеназ:
Каталаза содержится в крови, костном мозге, мембранах слизистых оболочек, почках и печени.
Во многих тканях, включая и печень, обнаружены микротельца − пероксисомы, которые богаты аэробными дегидрогеназами и каталазой. Биологически выгодно группировать в одном месте как ферменты, приводящие к образованию Н2О2, так и ферменты, разлагающие это соединение.
К ферментам, катализирующим образование Н2О2, кроме пероксисомных ферментов, относятся также митохондриальные и микросомные системы транспорта электронов.
Оксигеназы.Ферменты этой группы катализируют включение кисло-рода в молекулу субстрата, которое происходит в две стадии:
1) кислород связывается с активным центром фермента;
2) происходит реакция, в результате которой, связанный кислород восстанавливается или переносится на субстрат.
Оксигеназы не относятся к ферментам, которые катализируют реакции, снабжающие клетку энергией; они участвуют в синтезе и деградации многих типов метаболитов, токсинов и ксенобиотиков. Оксигеназы подразделяются на 2 подгруппы.
Монооксигеназы (гидроксилазы). Эти ферменты катализируют включение в субстрат только одного из атомов молекулы кислорода. Другой атом кислорода восстанавливается до воды; для чего необходим дополнительный донор электронов (косубстрат):
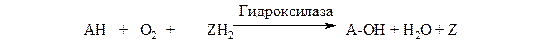
субстрат косубстрат
Природа косубстрата может быть различной. Многие оксигеназы используют НАДФН2. В этом случае суммарное уравнение реакции можно записать как
 К подобным системам относятся микросомные цитохром-Р-450-содержащие монооксигеназные системы, участвующие в метаболизме многих лекарственных веществ путем их гидроксилирования. Они находятся в микросомах печени вместе с цитохромом Р-450 и цитохромом b-5. Восстановителями этих цитохромов являются НАДН и НАДФН. Цитохромы окисляются субстратами в результате серии ферментативных реакций, составляющих так называемый гидроксилазный цикл:
К подобным системам относятся микросомные цитохром-Р-450-содержащие монооксигеназные системы, участвующие в метаболизме многих лекарственных веществ путем их гидроксилирования. Они находятся в микросомах печени вместе с цитохромом Р-450 и цитохромом b-5. Восстановителями этих цитохромов являются НАДН и НАДФН. Цитохромы окисляются субстратами в результате серии ферментативных реакций, составляющих так называемый гидроксилазный цикл:
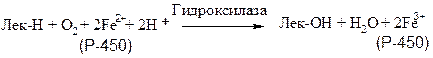
Лек - лекарственное вещество
К лекарственным веществам, метаболизм которых идет при участии рассматриваемых систем, относятся, например, морфин и бензпирен. Многие лекарственные вещества (например, фенобарбитал) способны индуцировать синтез микросомных ферментов и цитохрома Р-450. Цитохром Р-450 является одновременно гемо- и флавопротеином, причем флавиновые нуклеотиды выступают в качестве второго донора электронов. Цитохром Р-450 в печени осуществляет гидроксилирование липофильных соединений, образующихся в качестве побочных продуктов или попадающих в организм извне (такие вещества объединяют под общим названием «ксенобиотики»). Введение в их молекулы гидроксильных групп повышает гидрофильность этих веществ, понижает токсичность и облегчает их вывод из организма.
Другой пример подобных систем − митохондриальные цитохром-Р-450-содержащие монооксигеназные системы. Они находятся в коре надпочечников, семенниках, яичниках, плаценте, почках и участвуют в биосинтезе стероидных гормонов из холестерина, катализируют гидроксилирование гидроксихолекальциферола, что приводит к образованию активной формы витамина D3 (1,25-дигидроксихолекальциферола). В коре надпочечников содержание митохондриального цитохрома Р-450 в 6 раз выше, чем содержание цитохромов в дыхательной цепи.
Диоксигеназы (истинные оксигеназы). Эти ферменты катализируют включение в молекулу субстрата обоих атомов молекулы кислорода:
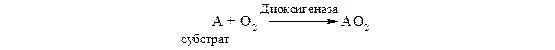
Примером служат железосодержащие ферменты 1.13.11.5 гомогентизатдиоксигеназа и 1.13.11.6 3-гидроксиантранилатдиоксигеназа, а также некоторые гемсодержащие ферменты, в частности триптофандиоксигеназа (триптофанпирролаза).
II. Трансферазы
Трансферазы катализируют реакции межмолекулярного переноса различных атомов, групп атомов и радикалов (кроме водорода). Деление на подклассы определяется природой переносимых групп.
Трансферазы, катализирующие реакции переноса одноуглеродных остатков. Например, фермент 2.1.2.1 серингидроксиметилтрансфераза, катализирующий образование метилентетрагидрофолиевой кислоты:
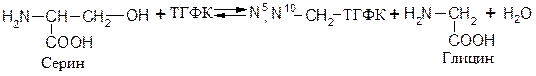
Ацилтрансферазы, катализирующие перенос ацильных групп. Например, фермент 2.3.1.21. карнитин-ацилтрансфераза, катализирующий перенос ацильной группы с атома серы коэнзима А (КоА) на гидроксильную группу карнитина:
Ацил-SКоА + Карнитин ® Ацилкарнитин +НSКоА
Гликозилтрансферазы, катализирующие перенос углеводных остатков. К этому подклассу относится фермент 2.4.1.1 гликогенфосфорилаза, участвующий в распаде резервного полисахарида − гликогена:
(С6Н10О5)n + Н3РО4 ® глюкозо-1-фосфат + (С6Н10О5)(n-1)
Гликоген Гликоген
Аминотрансферазы (трансаминазы), катализирующие перенос аминогруппы в реакциях трансаминирования. Трансаминазы в качестве кофермента используют пиридоксальфосфат. Примером может служить фермент 2.6.1.2 аланинаминотрансфераза, катализирующий реакцию трансами
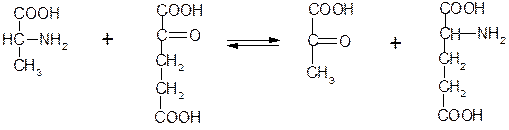 |
нирования между аланином и a-кетоглутаратом:
Аланин a-Кетоглутарат Пируват Глутамат
Фосфотрансферазы (киназы), катализирующие перенос остатка фосфорной кислоты от нуклеозидтрифосфатов. Например, фермент 2.7.1.1 гексокиназа , катализирующий следующую реакцию:
D-гексоза + АТФ ® D-гексозо-6-фосфат + АДФ
Гексокиназа обладает групповой субстратной специфичностью и способна фосфорилировать различные гексозы: глюкозу, маннозу, галактозу и др.
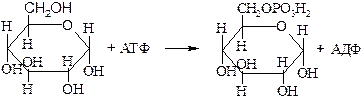 |
Глюкоза Глюкозо-6-фосфат
III. Гидролазы
Гидролазы катализируют реакции гидролиза. Подклассы определяются в соответствии с типом связей, подвергающихся гидролизу.
Эстеразы, катализирующие гидролиз сложноэфирных связей. Например, панкреатическая липаза, которая катализирует гидролиз сложноэфирных связей в жирах.
Пептидазы (протеазы), катализирующие гидролитическое расщепление пептидных связей в белках и пептидах. Например, ферменты пищеварительного тракта: 3.4.21.1 химотрипсин, 3.4.21.4 трипсин, 3.4.23.1 пепсин, 3.4.23.15 ренин.
Гликозидазы, катализирующие гидролиз гликозидных связей в полисахаридах. Например, 3.2.1.1 a-амилаза, которая катализирует гидролиз a(1→4)-О-гликозидных связей в полисахаридах.
IV. Лиазы
Лиазы катализируют реакции негидролитического расщепления связей С−С, С−О, С−N (часто с образованием двойных связей), а также реакции образования новых связей без участия энергии, выделяемой при гидролизе АТФ.
Декарбоксилазы, катализирующие реакции удаления углекислого газа из субстрата. Их коферментом является пиридоксальфосфат. Например, фермент 4.1.1.15 глутаматдекарбоксилаза, катализирующий образование тормозного нейромедиатора g-аминомасляной кислоты:
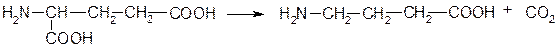
Глутаминовая кислота g-Аминомасляная кислота (ГАМК)
Гидролиазы, катализирующие обратимое отщепление воды с образованием двойных связей. Например, фермент 4.2.1.2 малатгидролиаза (фумараза):
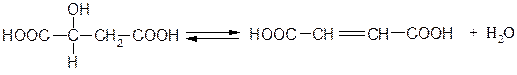
Малат Фумарат
V. Изомеразы
Изомеразы катализируют реакции внутримолекулярного переноса групп, а также взаимопревращение геометрических и оптических изомеров.
Внутримолекулярные оксидоредуктазы, катализирующие взаимное превращение альдоз и кетоз. Например, фермент 5.3.1.1 D-глицеральдегид-3-фосфат-кетолизомераза (триозофосфатизомераза), который катализирует обратимое превращение глицеральдегид-3-фосфата и диоксиацетонфосфата:

Глицеральдегид-3-фосфат Диоксиацетонфосфат
Внутримолекулярные трансферазы (мутазы), катализирующие перемещение групп атомов внутри одной молекулы. Например, фермент 5.4.2.2 фосфоглюкомутаза, катализирующий обратимое превращение изомеров:
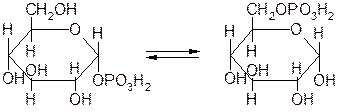
Глюкозо-1-фосфат Глюкозо-6-фосфат
Рацемазы и эпимеразы, катализирующие превращения энантиомеров и эпимеров соответственно. Например, фермент 5.1.3.2 УДФ-глюкозо-4-эпимераза, катализирующий взаимное превращение эпимеров УДФ-глюкозы и УДФ-галактозы:
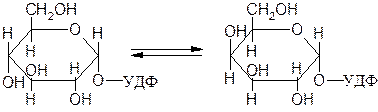
Уридиндифосфатглюкоза Уридиндифосфатгалактоза
VI. Лигазы (синтетазы)
Лигазы катализируют реакции образования из более простых молекул более сложных с использованием энергии гидролиза АТФ.
Ферменты, катализирующие образование связей С−N. Например, фермент 6.3.1.2 глутаминсинтетаза, катализирующий образование амида глутаминовый кислоты:
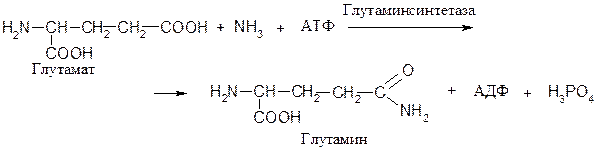
Ферменты, катализирующие образование связей С−С. Например, фермент 6.4.1.1 пируват: двуокись углерода-лигаза (пируваткарбоксилаза), при участии которого происходит превращение пирувата в оксалоацетат:

Пируват Оксалоацетат
|
из
5.00
|
Обсуждение в статье: VI. Лигазы (синтетазы) |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы