 |
Главная |


Рибонуклеиновые кислоты
|
из
5.00
|
Гетерогенность молекул РНК.Содержащиеся в клетке РНК различаются размером, составом, функциями и локализацией. В цитоплазме содержится стабильная РНК нескольких видов: транспортная РНК (тРНК), матричная, или информационная (мРНК, или иРНК), рибосомная (рРНК). В ядре локализована ядерная РНК (яРНК), количестве которой составляет от 4 до 10% от суммарной клеточной РНК. В состав яРНК входит большое число молекул, различающихся по размерам и нуклеотидный последовательностям, существенно превышающее число различных молекул цитоплазматических мРНК.
Наряду с основными видами РНК из зараженных вирусами клеток можно выделить геномные РНК вирусов растений, некоторых вирусов бактерий (например, бактериофаг Q (3 кишечной палочки) и некоторых вирусов животных (вирус полиомиелита). Геномные РНК хранят и передают следующему поколению соответствующую генетическую информацию. Они относятся к самым крупным: их молекулярная масса достигает нескольких миллионов, а число нуклеотидов — десятков тысяч.
Молекула РНК в отличие от ДНК состоит (за редким исключением) из одной полинуклеотидной цепи. Полинуклеотидная цепь РНК, закручиваясь на себя, образует в палиндромных участках короткие двухспиральные «шпильки», в которых азотистые основания образуют комплементарные пары: Г с Ц, А с У. Это довольно прочные структуры, которые видны под электронным микроскопом.
Транспортная РНК.Транспортные РНК — самые мелкие молекулы РНК. Они включают в себя от 75 до 90 нуклеотидных единиц, М-23 000—30 000. тРНК составляют 10—20% суммарной РНК клетки. Их функция состоит в том, чтобы транспортировать аминокислоты в рибосомы и ставить их в определенные участки полипептидной цепи при ее биосинтезе. Таким образом, тРНК участвует в процессе трансляции, причем играет роль адаптора, т. е. своеобразного переводчика: переводит последовательность нуклеотидов в последовательность аминокислотных остатков белковой молекулы. Каждой из 20 аминокислот соответствует своя тРНК. Для некоторых аминокислот известно несколько тРНК. Например, существует пять различных тРНК, переносящих лей, и пять различных тРНК, переносящих сер. В то же время каждый вид тРНК переносит в рибосому только один вид аминокислоты, «свою» аминокислоту. В клетке присутствует до 60 разных видов тРНК.Кроме четырех обычных рибонуклеотидов (А, Г, Ц и У) в тРНК содержится много (8—19%) минорных нуклеотидов. Список минорных компонентов тРНК включает до 60 названий: различные метилированные аденины и гуанины, метилированные пиримидины (тимин,5-метилцитозин) и др. Не все они встречаются в какой-либо одной молекуле тРНК, но универсальными и наиболее распространенными являются псевдоуридин и дигидроуридин.
Молекула тРНК представляет собой одиночную полинуклеотидную цепь, закрученную «на себя». Она образует сложную пространственную структуру.
Вторичная структура тРНК характеризуется частичной спирализацией молекулы. В центре молекулы находится неспирализованная область. 3'- и 5'-Концы полинуклеотидной цепи спарены, образуют акцептирующий стебель. Это самый длинный спирализованный участок (7 пар). Он завершается на З'-конце в большинстве случаев неспаренной последовательностью ЦЦА. К 3'- или 2/-ОН-группе концевого аденозинового остатка присоединяется соответствующая аминокислота через свою СООН-группу, образуется аминоацил-тРНК.

Рис. 7. Структура транспортных РНК:/ — общая схема строения, // — третичная структура
Метилированные и другие модифицированные нуклеотиды располагаются в тРНК в участках, не вовлеченных в образование водородных связей. Возможно, они играют некоторую роль в образовании третичной структуры тРНК.
Матричная (информационная) РНК. Матричная РНК образуется в процессе транскрипции. Она несет точную копию генетической информации, закодированной в определенном участке ДНК, а именно информации о последовательности аминокислот в белках. У прокариот матричные РНК (мРНК) образуются сразу в процессе транскрипции. В эукариотических клетках в процессе транскрипции вначале образуются про-мРНК. Затем протекает процессинг, в ходе которого первичные транскрипты превращаются в мРНК. Свое название матричная РНК получила в связи с той функцией, которую она выполняет в клетке: она служит матрицей, на которой синтезируется полипептидная цепь в рибосоме. Каждой аминокислоте соответствует в мРНК определенная тройка (триплет) нуклеотидов, называемая кодоном этой аминокислоты. Последовательность кодонов в цепи мРНК определяет последовательность аминокислот в белке. Поскольку мРНК несет наследственную информацию о первичной структуре белка, нередко ее называют информационной РНК (иРНК).
Структура рибосом и рибосомной РНК. Рибосомная (рибосомальная) РНК (рРНК) — это та основа, на которой располагаются белки, образуя рибосому. На электронных микрофотографиях рибосомы видны как плотные округлые гранулы приблизительно сферической формы. Число рибосом в клетке очень велико: у бактерий в среднем 104, в эукариотических клетках— 106. Рибосомы локализуются главным образом в цитоплазме, кроме того,— в ядре (особенно в ядрышке), митохондриях и хлоропластах.
По размерам и молекулярной массе все изученные до сих пор рибосомы делят на три группы. Первую группу образуют относительно мелкие (30x30x20 нм) бактериальные рибосомы.
Вторую группу образуют крупные (40×40×20 нм) рибосомы эукариотических клеток. Они имеют константу седиментации 80S и М≈4,5х106. Как и рибосомы первой группы, они состоят из двух субчастиц. Малая 405-субчастица содержит 18S РНК и 30 белков. Большая 60S-субчастица содержит 28S РНК, 5S РНК и 5,8S РНК, а также 41 белок (табл. 8).
Таблица 8. Размеры рибосомных РНК
| Объект исследования | Константа седиментации, S | Молекулярная масса | Число нуклеотидов |
| E.coli Печень крысы | 5,8 | 1,1 ×106 0,56 ×106 4,1 ×104 1,7 ×106 0,65 ×106 5 ×104 4 ×104 | 121 |
Третью группу составляют рибосомы митохондрий и хлоропластов эукариотических клеток. Рибосомы митохондрий в общем относятся к классу 70S. Однако они различаются по коэффициентам седиментации у разных групп эукариот. Так, у грибов и эвгленовых коэффициент седиментации митохондриальных рибосом составляет 70—74S, у высших животных — 55—60S, у высших растений — около 80S. Рибосомы хлоропластов, напротив, более однородны по этому признаку, коэффициент их седиментации равен 67—70S.
При синтезе белка определенное число рибосом (от 3 до 80— 100) прикрепляется к длинным нитевидным молекулам мРНК, образуя полисомы. Каждая рибосома в полисоме
способна синтезировать полную полипептидную цепь. Образование групп рибосом повышает эффективность использования мРНК, поскольку на ней может одновременно синтезироваться несколько идентичных полипептидных цепей. Полисомы находятся или в свободном состоянии, или в тесной связи с мембранами эндоплазматической сети. мРНК, кодирующие внутриклеточные белки, содержатся преимущественно в свободных полисомах, а мРНК, кодирующие секреторные белки,— в мембраносвязанных.
Биосинтез нуклеотидов
Пути синтеза мононуклеотидов. Пуриновые и пиримидиновые нуклеотиды могут синтезироваться de novo, т. е. из простых предшественников, а также непосредственно из готовых пуриновых и пиримидиновых оснований. Относительное значение этих двух путей существенно отличается для разных клеток. Так, в тканях млекопитающих нуклеотиды преимущественно синтезируются de novo, хотя в быстро растущих тканях образуются обоими путями. Напротив, для нормального роста и развития многих видов бактерий необходимо наличие в питательной среде готовых молекул пуринов или пиримидинов.
Биосинтез пуриновых нуклеотидов. Биосинтез пуринового кольца de novo протекает одинаково у разных видов живых существ: у бактерий, дрожжей, птиц, человека. Это один из многих примеров единства ряда основных биохимических процессов всего живого. Эксперименты с использованием изотопов позволили установить, из какого предшественника поступает каждый атом пуринового кольца (рис. 8).
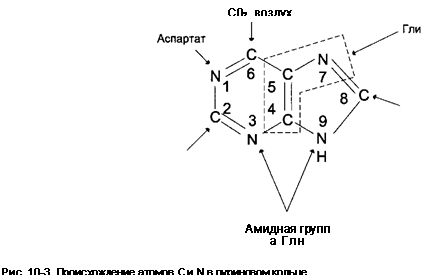
Рис. 8. Происхождение атомов пуринового кольца
Таким образом, синтезируется не свободное азотистое основание, а его нуклеотид — инозиновая кислота. Инозиновая кислота служит предшественником всех остальных пуриновых нуклеотидов. Так, адениловая кислота (АМФ) образуется в результате аминирования ИМФ, аминогруппа поставляется аспарагиновой кислотой. Образование гуаниловой кислоты (ГМФ) из ИМФ является двухстадийной реакцией. Первая стадия состоит в окислении ИМФ до ксантозин-5'-фосфата (КМФ). Затем происходит его аминирование, и образуется ГМФ. Донор аминогруппы, повидимому, различен у ряда организмов: у птиц и млекопитающих донором служит глицин, в бактериальных системах — NH3.
Биосинтез пиримидиновых нуклеотидов. Применение метода изотопов позволило выяснить происхождение отдельных звеньев пиримидинового ядра.
Первым этапом в синтезе пиримидиновых нуклеотидов является образование карбамоилфосфата из NH3 и СО2. Карбамоилфосфат вступает в реакцию с аспаргин и под действием аспартат-карбамоилтрансферазы превращается в карбамоиласпартат. Последний подвергается циклизации и окислению, в результате чего образуется оротовая кислота, т. е. завершается формирование пиримидинового кольца.
Превращение урацила в цитозин происходит на уровне нуклеозидтрифосфатов. Оно осуществляется под действием фермента ЦТФ-синтетазы: УТФ + NН3+АТФ→ЦТФ+АДФ + Фн. Тиминовые нуклеотиды образуются в результате метилирования дезоксиуридинмонофосфата. Реакция катализируется ферментной системой, которую часто называют тимидилат-синтазой. Процесс весьма сложен и протекает в несколько стадий. Источником одноуглеродного фрагмента при С-5 служит кофермент N5,N10-метилентетрагидрофолиевая кислота.
Биосинтез дезоксирибонуклеотидов. Превращение рибозы в дезоксирибозу происходит на нуклеотидном уровне. Механизм этого процесса был выяснен при изучении восстановления рибонуклеотидов в экстрактах Е. coli. Фермент рибонуклеозид-дифосфатредуктаза катализирует восстановление всех четырех рибонуклеозиддифосфатов — АДФ, ГДФ, ЦДФ, УДФ в их дезоксипроизводные дАДФ, дГДФ, дЦДФ, дУДФ. Источник восстанавливающей способности фермента in vivo неизвестен.
In vitro активны два донора водорода. Один из них — низкомолекулярный серосодержащий белок тиоредоскин, две SH-группы которого окисляются с образованием дисульфидного мостика. Вторым донором водорода может служить восстановленный глутатион. Редуктазные системы, выделенные из различных животных клеток, сходны с описанной системой Е. coli.
|
из
5.00
|
Обсуждение в статье: Рибонуклеиновые кислоты |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы