 |
Главная |


Экспериментальные доказательства модели Уотсона – Крика
|
из
5.00
|
1953 г. считается годом рождения новой науки – молекулярной биологии.
Конкретные доказательства того, что ДНК двойная спираль, были получены через 5 лет в опытах Метью Мезельсона и Франклин Сталя.
Стент предложил проверить 3 варианта репликации ДНК.
1. Консервативный – новые молекулы не содержат материала родительской ДНК. Синтез идет как при полуконсервативном типе, но затем две вновь синтезированные цепи объединяются в одну молекулу, две родительские реассоциируют в «консервативную» первоначальную молекулу.
2. Полуконсервативный – каждая дочерняя молекула ДНК состоит из одной родительской и одной вновь синтезированной цепей.
3. Дисперсионный – родительская двойная спираль раскручивается с разрывом цепей при каждом полуобороте. Вдоль фрагментов происходит синтез, а затем все эти фрагменты соединяются крест-накрест и образуют две новые спирали. В этом случае обе цепи каждой двойной оказываются составленными из чередующихся старых и новых фрагментов.
Для определения способа репликации ДНК, необходимо уметь отличить дочерние молекулы от родительских.
Исследователи выращивали E. coli на среде c N15 (15NH4Cl) в течение двенадцати поколений. Тяжелый изотоп N15 включается в состав ДНК и служит меткой. Молекулы ДНК, содержащие легкий 14N и тяжелый 15N, отличаются по плотности и могут быть разделены высокоскоростным равновесным центрифугированием в градиенте плотности хлористого цезия, собираясь в том слое, плотность которого равна плотности молекул.
Плотность ДНК клеток E. coli, содержащей тяжелый 15N, 1,724 г/см. плотность ДНК клеток E. coli, содержащей легкий 14N, 1,710 г/см. Смесь этих двух типов ДНК легко разделяется при центрифугировании по плотности.
Бактерии, которые культивировались на среде с 15N, переносились на среду14N.
Результаты:
1. после первого деления клеток плотность ДНК была промежуточной.
2. после второго деления клеток на той же среде половина клеток имела легкую ДНК, а вторая половина – промежуточную.
3. после третьего деления на среде с 14N 3/4 ДНК имело плотность, равную плотности легкой ДНК и ¼ сохраняло промежуточную плотность.
4. Таким образом, между числом генераций и распределением плотности была прямая связь, что в точности соответствовало полуконсервативному типу репликации ДНК, предсказываемому моделью Уотсона – Крика. Было показано, что ДНК с промежуточной плотностью – это гибридная двойная спираль, одна из цепей которых содержит только тяжелый изотоп азота 15N, а другая – только легкий 14N. При нагревании ДНК промежуточной плотности в течение 30 мин при +1000 С образовались две равные по объему фракции ДНК с разными плотностями, одна из которых имела плотность тяжелой ДНК, а другая – плотность легкой ДНК.
Полуконсервативный механизм репликации был доказан на высших растениях – на конских бобах, Vicia faba, в 1957 г. – Дж. Тейлор, Филипп Вудс и Вальтер Хьюгес. Они исследовали хромосомы в быстро деляшихся клетках корневой меристемы, набдюдали включение 3Н-тимидина, меченного тритием.
Генетический код
Информация о синтезе белков заключена в ДНК в кодирующих участках. Первичная структура каждого белка в организме кодируется определенным геном.
Известно, что в состав белков входит 20 основных различных аминокислот, в то время как в ДНК содержатся нуклеотиды только четырех видов. Следовательно, соответствие между нуклеотидными и аминокислотными последовательностями не может быть построено по принципу «один к одному». Поэтому необходимо предположить существование такого кода, в котором каждой данной аминокислоте соответствовала бы комбинация из нескольких нуклеотидов.
Последовательность нуклеотидов в молекуле ДНК, опеределяющая последовательность аминокислот в белке, называетсягенетическим кодом(соответствие меду нуклеотидными триплетами и кодируемыми ими аминокислотами).
Концепция о генетическом коде очень важна, из нее, в частности, вытекает представление о существовании системы передачи наследственной информации. «Генетический код, по словам Крика, устанавливает связь между двумя великими полимерными языками – языком нуклеиновых кислот и языком белков».
Особенности генетического кода:
1. Триплетный – одна аминокислота кодируется последовательностью из трех нуклеотидов, называемой кодоном.
На основе триплетов можно составить 64 кодона = 43, из них только 61 используется для кодирования аминокислот (смысловые кодоны). Один из них стартовый – метиониновый.
2. Недвусмысленный,то естьоднозначно идентифицированный, так как каждый триплет кодирует только одну аминокислоту.
3. Вырожденный.
Поскольку 20 аминокислот кодируется 61 триплетом, то, очевидно, должны быть кодоны «синонимы», то есть различные триплеты, кодирующие одну и ту же аминокислоту.
Код, в котором одной аминокислоте соответствует более чем один кодон, называется вырожденным. Лейцин, например, кодируется шестью кодонами. Кодоны-синонимы одной аминокислоты размещены в таблице неслучайно, а почти всегда находятся в одном и том же квадрате и различаются друг от друга только по последнему нуклеотиду. Исключение составляют лейцин, серин, аргинин, кодирующиеся шестью кодонами и не размещающиеся в одном квадрате. Синонимичные кодоны используются различными организмами с разной частотой. Из четырех кодонов для глицина GGA используется в структурных генах человека в 26% случаев, а в Escherichia coli — в 9%. Такая же ситуация наблюдается и для стоп-кодонов. Так, у человека частота использования кодонов UAA, UAG и UGA составляет 0,22, 0,17 и 0,61 соответственно, а у E. coli — 0,62,0,09 и 0,30. Несмотря на все эти различия, генетический код у всех организмов, за редким исключением, одинаков.
3. Код содержит три бессмысленных кодона ( нонсес кодоны, стоп-кодоны – УАА. УАГ, УГА), не кодирующие ни какую аминокислоту.
4. Непрерывный.С началом транскрипции кодоны мРНК считываются один за другим без перерывов.
5.Неперекрывающийся.
Каждой аминокислоте в полинуклеотидной цепи соответствует отдельная тройка нуклеотидов. Каждое основание может принадлежать только одному триплету и не является частью другого триплета. Кодоны ни чем не отделены друг от друга. Код считывается с фиксированной точки в пределах гена в одном направлении.
6. Универсален.
Он одинаков у всех организмов: одни и те же триплеты кодируют одну и ту же аминокислоту у всех организмов. Отличие - митохондриальный генетический код имеет отличия от универсального кода. Но при этом он одинаков у всех позвоночных, но отличается от дрожжей и дрозофил.
Экспериментальное определение смысла каждого кодона можно отнести к самым выдающимся достижениям современной науки. Ниренбергом и Матеи в 1961 г. был расшифрован триплет УУУ, являющийся кодоном для фенилаланина. Прямой метод анализа, основанный на синтезе полипептидной цепочке in vitro на матрице синтетической РНК с известным соотношением случайно расположенных нуклеотидов. Полностью расшифровка генетического кода была завершена в 1966 г.
Корана (химик-органик) осуществил синтез всех 64 триплетов и оказалось возможным определить значение всех триплетов генетического кода.
Синтез белка
В молекуле нуклеиновой кислоты различают кодирующие – кодируют белки и некодирующие последовательности – регуляторные элементы. Подавляющее большинство генов содержит в закодированном виде информацию о синтезе белков. Белки — это биологические молекулы, участвующие практически во всех процессах, протекающих в живых системах. Они служат катализаторами разнообразных биохимических ре акции, осуществляют транспорт веществ внутри клеток и между клетками, регулируют проницаемость клеточных мембран, из них строятся различные структурные элементы. Белки участвуют в осуществлении двигательных функций, обеспечивают защиту от инфекций и токсинов, регулируют синтез остальных генных продуктов.
Основой структуры белка является полимерная цепочка, состоящая из мономеров-аминокислот – полипептид, аминокислотная последовательность. Основной структурной единицей белков являются аминокислоты. Все аминокислоты имеют сходное химическое строение, К центральному атому углерода (α-углерод) присоединены атом водорода (Н), аминогруппа (NH3+), карбоксильная группа (СОО—) и R-группа (боковая цепь) (рис. 3.7, А). Существует 20 разных боковых групп и соответственно 20 аминокислот.

Пептидная связь – связь между аминокислотами в полипептидной цепочке за счет взаимодействия аминогруппы одной аминокислоты с карбоксильной группой другой аминокислоты.
Информация о синтезе белков заключена в ДНК, которая почти вся находится в ядре – в кодирующих участках. В начале 50-х годов было показано, что синтез белка фактически происходит в цитоплазме и в нем участвуют рибосомы. Стало ясно, что должен существовать какой-то механизм, переносящий генетическую информацию из ядра в цитоплазму. В 1961 г. Франсуа Жакоб и Жак Моно предположили существование матричной РНК (мРНК).
Для синтеза белка необходимы следующие компоненты – м-РНК, т-РНК, р-РНК, аминокислоты и набор различных ферментов.
Переписывание генетической информации, записанной в нуклеотидной последовательности ДНК, в нуклеотидную последовательность РНК, называется транскрипцией.
В зоне синтеза РНК происходит расплетание спирали ДНК и считывается информация с участка цепи ДНК-матрицы, кодируется только одна цепь молекулы ДНК. В процессе синтеза макроэнергетические связи разрываются и их энергия используется на построение молекулы РНК.
На ДНК-матрице транскрибируется РНК трех типов – мРНК, рРНК и тРНК, участвующие в передаче информации от РНК к белку. У большинства прокариот транскрипция всех РНК осуществляется с помощью одной и той же РНК-полимеразы. У эукариот мРНК, рРНК и тРНК транскрибируются разными РНК-полимеразами.
Транскрипционные единицы ограничиваются промоторами – участок инициации и терминаторами. Транскрипция осуществляется с помощью фермента ДНК-зависимая-РНК-полимераза, который связывает нуклеотиды между собой, образуя полинуклеотидную цепь и-РНК – служит матрицей для построения белковой цепи (3 – 5 % всей РНК в клетке).
С молекулярной точки зрения ген представляет собой специфическую нуклеотидную последовательность, транскрибируемую в РНК. Подавляющее большинство транскрибируемых последовательностей ДНК составляют так называемые структурные гены, на которых синтезируются мРНК. Каждая последовательность из трех оснований в молекуле и-РНК называется кодоном. Конечным продуктом структурного гена является белок.
У прокариот структурный ген представляет собой непрерывный участок молекулы ДНК. Транскрипция начинается со связывания РНК-полимеразы с промотором, и далее последовательно копируется весь структурный ген (кодирующая область) от первого нуклеотида до последнего с образованием функциональной мРНК.
У эукариот большинство структурных генов состоит из нескольких дискретных кодирующих областей (экзонов), разделенных некодирующими областями (интронами). По завершении транскрипции эукариотического структурного гена интроны вырезаются из первичного продукта транскрипции с помощью ферментов, а экзоны сшиваются друг с другом «торец в торец» (сплайсинг) с образованием функциональной мРНК (рис. 1 и рис. 2). Обычно длина экзонов составляет от 150 до 200 нуклеотидов, а длина интронов варьирует от 40 до 10 000 нуклеотидов. Очень немногие эукариотические структурные гены вообще не имеют нитронов. Иногда сплайсинг мРНК может проходить по альтернативному варианту. Например, в одной ткани функциональная мРНК может образовываться в результате соединения всех экзонов первичного транскрипта, а в другой какой-то экзон будет вырезан вместе с фланкирующими его интронами и образуется другая функциональная мРНК. Благодаря альтернативному сплайсингу в разных тканях могут образовываться разные продукты одного и того же структурного гена.

Рис 1. Схематическое изображение прокариотического структурного гена
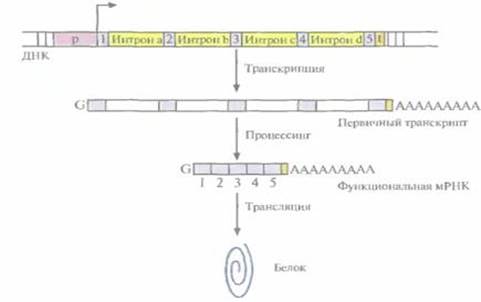
Рис 2. Схематическое изображение эукариотического структурного гена.
Синтезированные молекулы и-РНК выходят из ядра через ядерные поры и направляются к рибосомам.
Синтез белка происходит в рибосомах. Рибосомы – это компактные рибонуклеиновые частицы, построенные из двух субчастиц, каждая из которых построена из нескольких белков, связанных с одной молекулой РНК – рибосомной РНК – р-РНК (составляет 80 % всей РНК клетки). При понижении концентрации ионов Мg рибосомы диссоциируют на субчастицы.
Бактериальные рибосомы сидементируют обычно при 70 S, а отдельные субчастицы при 50 S и 30 S. Большая субчастица имеет почти сферическую форму и по размеру примерно вдвое больше маленькой асимметричной субчастицы.
Цитоплазматические рибосомы высших эукариот больше бактериальных и обычно сидементируют при 80 S. Скорость сидементации отдельных субчастиц составляет 60 S и 40 S (S – константа Сведберга, используемая в качестве единицы размера, который оценивают по скорости сидементации частиц в растворе при центрифугировании).
Хлоропласты и митохондрии эукариотических клеток располагают своими собственными рибосомами, которые по размеру ближе к прокариотическим, чем к цитоплазматическим рибосомам эукариот. Большая часть ДНК, обнаруживаемой в хлоропластах и митохондриях, содержит информацию о структуре компонентов их собственной системы биосинтеза белка.
Синтез всех полипептидных цепей протекает в направлении от N-концак С-концу и всегда начинается с метионина.
Трансляция (от translation – перевод) – процесс перевода информации, заключенной в последовательности нуклеотидов в и-РНК, в последовательность аминокислот белка. В процессе трансляции принимает участие м- РНК, т- РНК (16 %), р- РНК (80 % всей РНК клетки) .
Каким образом происходит трансляция?
Информация об аминокислотной последовательности каждого белка кодируется в виде последовательности кодонов в соответствующей м-РНК.
Строение аминокислот и нуклеотидных триплетов не является комплементарным, как у двух цепочек ДНК или даже ДНК и РНК, и прямое перекодирование оказывается невозможным. Кодоны и аминокислоты не вступают в непосредственное взаимодействие друг с другом, Связь между ними осуществляется при участии молекул т- РНК. Транспортная РНК играет роль «адаптора» и выполняет сразу две функции: узнает кодон и соответствующую аминокислоту. Нуклеотидную последовательность (типичные т-РНК состоят из 75-85 нуклеотидов, растворимая низкомолекулярная РНК) можно представить в видеконфигурации «клеверного листа». Участки молекулы, комплементарные друг другу, спариваются и образуют «стебли», а одноцепочечные участки остаются в виде петель. Часть молекулы, состоящую из стебля и петли, называют «шпилькой». В нижней петле расположен триплет, комплементарный кодону той аминокислоты, которую представляет т-РНК – антикодон – последовательность из трех нуклеотидов.Поэтому т-РНК узнает кодон путем комплементарного спаривания оснований. На 3-конце тРНК находится последовательность рЦрЦрАон, которая служит специфическим сайтом ковалентного присоединения соответствующей аминокислоты (акцепторный конец).
|

Для каждой из 20 аминокислот существует не менее одного вида специализированной тРНК. Последовательность нуклеотидов, идущих после аденозин-цитозинового окончания, у различных типов т-РНК различна: этим достигается их специфичность. Различные транспортные РНК отличаются друг от друга как по нуклеотидной последовательности, так и по содержанию некоторых необычных (минорных) оснований. Этим достигается специфичность тРНК.
Ковалентное связывание аминокислоты с соответствующей РНК происходит при участии специфического фермента аминоацил-т РНКсинтетазы. Существует 20 видов ферментов этого типа. Связывание аминокислоты с т РНК протекает в две стадии:
1. Активация аминокислот. Аминокислота взаимодействует с АТФ под контролем аминоацил-тРНК-синтетазы, образуя при этом аминоациладенилаты. Каждый фермент специфичен для одной аминокислоты.
2. Перенос активированной аминокислоты на 3-ОН-конец соответствующей тРНК, при этом образуется аминоацилированная тРНК.
Процесс синтез белка состоит из следующих этапов:
1. инициация;
2. элонгация;
3. терминация.
1. Инициация.
Рибосомы сами по себе не обладают активностью. Для включения в синтез белков они должны присоединиться к мРНК в присутсвии ионов Mg +2.
Инициация полипептидной цепи начинается с присоединения малой рибосомной субъединицы к соответствующему центру связывания на мРНК, включающему инициаторный метиониновый кодон АУГ. АУГ и ГУЦ кодоны инициации у бактерий, у эукариот только АУГ – инициирующая тРНК – тРНК, ее метионин не формилирован. Затем происходит взаимодействие комплекса с большой рибосомной субъединицей. В большой рибосомной субъединице находится два участка связывания т-РНК:
Р участок – донорный – участок связывания пептидил-т РНК;
А участок – акцепторный – участок связывания аминоацил- т РНК.
Пептидил-тРНК представляет собой тРНК, несущую целую полипептидную цепь, синтезированную к данному моменту. Инициация относительно медленная стадия синтеза белка.
2. Элонгация.
Включает все реакции от образования первой пептидной связи до присоединения последней аминокислоты, наиболее быстрая стадия (последовательное соединение аминокислот путем образования пептидных связей между аминокислотными остатками с одновременным освобождением молекул тРНК).
Следующий кодон, стоящий сразу после инициирующего АУГ кодона попадает в участок А, где может происходить взаимодействие с аминоацил-тРНК, несущей соответствующий антикодон, который будет комплементарен кодону. После связывания аминоацил-тРНК с А-участком происходит образование пептидной связи при участии фермента пептидилтрансферазы, входящего в состав большой рибосомной субъединицы.
При образовании пептидной связи происходит перенос остатка формилметионина ( fMet) на NH-группу следующего аминокислотного остатка, находящегося в виде аминоацил-тРНК в А-участке. При этом образуется пептидилтранспортная РНК.
Рибосома продвигается вдоль мРНК на расстояние, соответсвующее одному кодону. При этом деацилированная тРНК вытесняется из рибосомы, а удлинившаяся на одну аминокислоту пепетидил-тРНК поступает в участок Р, освобождая участок А. Освобожденный участок А занимает следующая аминоацил-тРНК, несущая антикодон, комплементарный кодону мРНК, находящемуся на данном этапе в участке А. Таким образом, синтез полипептида представляет собой многократное повторение цикла присоединения очередной аминокислоты к растущей полипептидной цепи.
По мере продвижения рибосомы вдоль мРНК, инициирующий участок цепи высвобождается и на нем происходит сборка следующего активного рибосомного комплекса, и на этой же матрице снова начинается синтез полипептида. При взаимодействии нескольких активных рибосом с единичной молекулой мРНК образуется полирибосома или полисома. Полисома – группа рибосом, расположенных вдоль одной молекулы мРНК. Например, при биосинтезе гемоглобина, состоящая из около 150 аминокислот, полисомная единица содержит в среднем около 5 рибосом на одну молекулу мРНК.
Синтез белка процесс быстрый. Время сборки цепей гемоглобина составляет 14 и 21 сек соответственно. Скорость синтеза белка зависит от температуры. У бактерий за 1 сек при t = 37 С включается от 12 до 17 аминокислот. Для синтеза среднего белка размером в 300 аминокислот требуется около 20 сек. В эукариотических клетках скорость белкового синтеза ниже, 2 аминокислоты в 1сек (в ретикулоцитах).
Терминация.
Полипептидный синтез по данной матрице продолжается до тех пор, пока в А-участке не окажется один из трех терминаторных кодонов мРНК – УАА, УАГ, УГА. Последовательность оснований узнается специализированными белковыми факторами терминации. В участок А подходит и терминирующий белок, вынуждающий пептидилтранферазу сделать «холостой» шаг. В результате этого законченная полипептидная цепь высвобождается и отделяется от рибосомы. Рисбосомные единицы в свою очередь диссоциируют, отделяются от мРНК и могут принять участие в синтезе следующей полипептидной цепи. Синтезированная полипептидная цепь приобретает свойственную ей вторичную, третичную или четвертичную структуру.
|
из
5.00
|
Обсуждение в статье: Экспериментальные доказательства модели Уотсона – Крика |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы