 |
Главная |


Регуляция амебоидной и мерцательной форм движения
|
из
5.00
|
Одним из свойств живой протоплазмы является сократимость, способность совершать движения, как одно из наиболее четких проявлений целесообразной саморегуляции в мире живых организмов. Различают два типа движения: движение протоплазмы внутри клетки и движение клетки по отношению к среде. Среди движений первого типа, можно выделить колебательное — наименее упорядоченное, хотя и наиболее широко распространенное, циркуляционное, ротационное (вращательное), фонтанирующее и направленное (в определенную сторону). Каждое из этих движений (в отдельности или в определенной комбинации) встречается в протоплазме клеток различных организмов, как одна из форм саморегуляции ее динамической структуры, и обеспечивает ряд дифференцированных внутриклеточных реакций на раздражение внешней -среды.
Второй тип движения обеспечивает активное изменение пространственного положения клетки относительно внешней среды. Различают четыре различные формы такого движения: амебоидное, ресничное, жгутиковое и мышечное. Наиболее простой формой механической работы является амебоидное движение. Хотя такое передвижение, естественно, происходит крайне медленно (средняя скорость составляет примерно 0,5—4,6 мк/с), оно лежит в основе различных приспособительных реакций примитивных живых организмов или специализированных клеток внутренней среды у млекопитающих. В основе амебоидного движения лежат вызываемые раздражающим фактором изменения обмена веществ, приводящие к местному изменению коллоидно-дисперсного состояния протоплазмы, к обратимому превращению геля в золь. Движущийся конец амебы разжижается, а сокращение геля «хвостовой части» этой клетки приводит к выпячиванию псевдоподии. В регуляции амебоидного движения значительная роль принадлежит химическому составу окружающей среды. Ионы кальция стимулируют, а ионы калия подавляют амебоидное движение. Дальнейшее совершенствование двигательной функции в историческом развитии мира живых существ обеспечивается формированием специальных морфологических структур с волокнистыми структурами — органелл движения — жгутиков и ресничек. Движения, совершаемые жгутиком или ресничкой, упорядочены. Реснички и жгутики выполняют разнообразные функции: загоняют органические частицы в органы пищеварения, вызывают движение воды в органах водного дыхания, осуществляют передвижения животного и т. д.
В простом случае мерцательного движения (маятниковые движения) реснички колеблются взад и вперед в одной плоскости, не меняя своей формы и лишь изгибаясь у своего основания. Более сложную форму представляет вращательное движение, а также свойственное более длинным жгутикам —движение, при котором по длине жгута проходят одновременно несколько колебательных волн (рис. 37), движение при помощи гребного удара и т. д.
Хотя абсолютная величина скорости перемещения, вызываемого ресничками, невелика (например, у парамеции 3 мм/с), но относительно размеров тела инфузории она весьма высока. При этом максимальная скорость движения (как и остановки движения) достигается у этих животных весьма быстро. Величина остаточного пробега клетки вычисляется по формуле

|
 где
где  масса носителя реснички;
масса носителя реснички;  —сопротивление при скорости движения
—сопротивление при скорости движения  .
.
Рис. 37. Волнообразное движение жгутика :
а — последовательные положения «стационарной» волны и б — бегущей волны; I—III — последовательные стадии процесса движения
Ритмичность сокращений ресничек определяется процессом, состоящим из двух этапов,— возбуждения, происходящего внутри реснички, и следующего за ним проведения механического изменения движения (рис. 38). Частота ударов определяется частотой
 Рис. 38. Предполагаемый механизм ритмического движения ресничек
Рис. 38. Предполагаемый механизм ритмического движения ресничек
|
сокращений и возбуждений одной реснички, играющей роль «водителя ритма». Возникающие импульсы распространяются на соседнюю ресничку и т. д.
Если рассматривать ресничку как жидкое тело, стремящееся благодаря поверхностному натяжению к максимальному уменьшению поверхности (принятию формы шара), можно принять следующую формулу для определения ее поверхностного натяжения:

где  —отношение радиуса шара к радиусу цилиндра;
—отношение радиуса шара к радиусу цилиндра;  —давление, необходимое для деформации;
—давление, необходимое для деформации;  —постоянная поверхностного натяжения;
—постоянная поверхностного натяжения;  —постоянная силы тяжести.
—постоянная силы тяжести.
Хотя в механизме мерцательного, а тем более амебоидного движения трудно различать структурные звенья механизма его регуляции, факты адаптивного поведения одноклеточных живых организмов являются достаточным основанием для утверждения наличия примитивного контроля движения уже на этой стадии развития живых существ.
Автоматическое движение ресничек одноклеточных и низших многоклеточных регулируется путем прямого воздействия внешней среды на реснитчатый аппарат. Автоматическая природа мерцательного движения сама по себе не может служить основанием для отрицания нервных влияний на него. Реснитчатые клетки ряда беспозвоночных способны менять интенсивность автоматического движения под влиянием притекающих нервных импульсов, причем эти импульсы носят исключительно задерживающий, тормозящий характер. Нервная регуляция у позвоночных животных представляет собой дальнейший прогресс в процессе регуляции движений; здесь наблюдаются не только тормозящий, но и ускоряющий, возбуждающий эффект (А. П. Шмагина, 1948).
Мышечное движение
Наиболее совершенной формой движения является сокращение мышечной ткани. Локомоторная функция организма обеспечивается своеобразными двигателями — мышцами. Подчеркивая своеобразие мышц как хемомеханических двигателей, В. С. Гурфинкель, С. В. Фомин (1973) отмечают следующие их особенности. Во-первых, мышца с функциональной точки зрения представляет собой набор большого числа параллельно (но не синхронно) работающих двигателей, несколько различающихся по своим механическим характеристикам. Во-вторых, мышца — это двигатель, в котором сократительная часть соединена последовательно с упругим элементом. Другая отличительная особенность мышцы — это выраженная нелинейность ее характеристик. В протоплазме мышечного волокна скелетной мускулатуры располагаются продольные волокнистые структуры — миофибриллы, представляющие структурную основу мышечного сокращения.
Последовательность процессов, лежащих в основе саморегуляции одиночного мышечного сокращения, имеет следующий вид. Возмущающий сигнал, поступающий к мышце,
воспринимается диском  или зоной соединений дисков А и
или зоной соединений дисков А и  при помощи поперечно ориентированной системы саркоплазматической сети. Вслед за этимпроисходит высвобождение ионов кальция, активация миофибрилл под действием АТФ, фиксация ионов кальция элементами саркоплазматической сети и выделение фактора расслабления, тормозящего действия АТФ-азы на миофибриллы.
при помощи поперечно ориентированной системы саркоплазматической сети. Вслед за этимпроисходит высвобождение ионов кальция, активация миофибрилл под действием АТФ, фиксация ионов кальция элементами саркоплазматической сети и выделение фактора расслабления, тормозящего действия АТФ-азы на миофибриллы.
 Одна из форм механической саморегуляции силы сокращения в зависимости от исходного растяжения хорошо проявилась в опытах, которые обнаружили гиперболическую зависимость изменения скорости изотонического сокращения скелетной мышцы при различной нагрузке. С увеличением длительности раздражения и частоты напряжение мышцы растет до некоторого максимума, который считают мерой способности сократительного механизма развивать напряжение. Как видно из графика (рис. 39), прирост напряжения увеличивается с натяжением линейно примерно до момента, когда достигается длина, соответствующая покоящейся мышце. При дальнейшем растяжении напряжение падает почти линейно, пока длина мышцы не достигает величины, примерно вдвое превышающей длину мышцы в состоянии покоя.
Одна из форм механической саморегуляции силы сокращения в зависимости от исходного растяжения хорошо проявилась в опытах, которые обнаружили гиперболическую зависимость изменения скорости изотонического сокращения скелетной мышцы при различной нагрузке. С увеличением длительности раздражения и частоты напряжение мышцы растет до некоторого максимума, который считают мерой способности сократительного механизма развивать напряжение. Как видно из графика (рис. 39), прирост напряжения увеличивается с натяжением линейно примерно до момента, когда достигается длина, соответствующая покоящейся мышце. При дальнейшем растяжении напряжение падает почти линейно, пока длина мышцы не достигает величины, примерно вдвое превышающей длину мышцы в состоянии покоя.
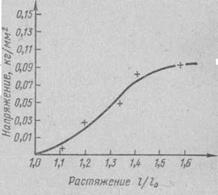
Кривая зависимости между напряжением и удлинением имеет S-образную форму (рис. 40).
 |
Скорость укорочения мышцы можно описать следующим уравнением:

|
где  —скорость укорочения в момент наблюдения;
—скорость укорочения в момент наблюдения;  —полное укорочение мышцы;
—полное укорочение мышцы;  —укорочение мышцы в данный момент.
—укорочение мышцы в данный момент.
Постоянная В определяет начальную скорость процесса. Одним из главных факторов, определяющих величину В, является нагрузка, приложенная к мышце (величина В уменьшается с ростом нагрузки).
Свойства сократительного механизма мышечной ткани обеспечивают возможность функционирования мышечных органов как систем с существенной нелинейностью в широком динамическом диапазоне (от минимального до оптимального режима). Выход на данный режим функционирования может служить еще одним рабочим механизмом саморегуляции мышечной деятельности, направленной на формирование двигательного акта, в то же время обеспечивая возврат к исходному состоянию при устранении возмущающего сигнала.
Сокращение сердечной мышцы является ярким примером нелинейности в биологических системах регуляции, оно подчиняется закону «все или ничего»: пороговые и надпороговые раздражения вызывают полное сокращение одинаковой амплитуды. Сила сокращения сердца зависит от степени растяжения его волокон, т. е. от степени кровенаполнения сердца (закон Старлинга). Такая реакция носит приспособительный характер и имеет важное значение для поддержания полноценного кровоснабжения. Так, переполненное кровью сердце растягивается, но благодаря усиливающимся сокращениям вся кровь выталкивается из него.
|
из
5.00
|
Обсуждение в статье: Регуляция амебоидной и мерцательной форм движения |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы