 |
Главная |


Фолликулярная/менструальная фаза
|
из
5.00
|
Репликация ДНК
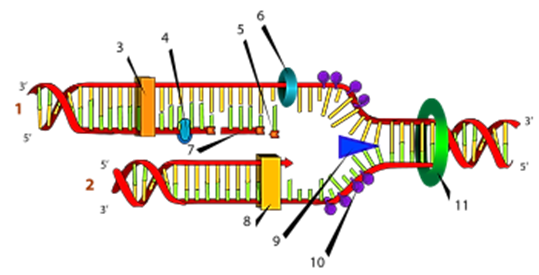
Схематическое изображение процесса репликации: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК-полимераза (Polα), (4) ДНК-лигаза, (5) РНК-праймер, (6) праймаза, (7) фрагмент Оказаки, (8) ДНК-полимераза (Polδ), (9) хеликаза, (10) одиночная нить со связанными белками, (11) топоизомераза.
Реплика́ция ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемый реплисомой.
Репликация проходит в три этапа:
- инициация репликации
- элонгация
- терминация репликации.
Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много.
Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта.
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК.
Суть репликации днк заключается в том, что специальный фермент разрывает слабые водородные связи, которые соединяют между собой нуклеотиды двух цепей. В результате цепи ДНК разъединяются, и из каждой цепи «торчат» свободные азотистые основания (возникновение так называемой вилки репликации). Особый фермент ДНК-полимераза начинает двигаться вдоль свободной цепи ДНК от 5'- к З'-концу (лидирующая цепь), помогая присоединиться свободным нуклеотидам, постоянно синтезируемым в клетке, к З'-концу вновь синтезируемой цепи ДНК. На второй нити ДНК (отстающая нить) новая ДНК образуется в виде небольших сегментов, состоящих из 1000—2000 нуклеотидов (фрагменты Оказаки).
Для начала репликации ДНК фрагментов этой нити требуется синтез коротких фрагментов РНК как затравок, для чего используется особый фермент — РНК-полимераза (праймаза). Впоследствии праймеры РНК удаляются, в образовавшиеся бреши встраивается ДНК с помощью ДНК полимеразы I. Таким образом, каждая цепь ДНК используется как матрица или шаблон для построения комплементарной цепи.
Основные ферменты репликации ДНК:
ДНК - полимераза
ДНК-полимераза - фермент, участвующий в репликации ДНК. Ферменты этого класса катализируют полимеризацию дезоксирибонуклеотидов вдоль цепочки нуклеотидов ДНК, которую фермент «читает» и использует в качестве шаблона. ДНК-полимераза начинает репликацию ДНК, связываясь с отрезком цепи нуклеотидов.
ДНК - лигазы
Лигаза - фермент, катализирующий соединение двух молекул с образованием новой химической связи (лиᴦᴎҏование). ДНК-лигазы -- ферменты, катализирующие ковалентное сшивание цеᴨȇй ДНК при репликации.
ДНК - хеликазы
ДНК хеликазы - ферменты раскручивающие двуцепочечную спираль ДНК.
ДНК-топоизомеразы
ДНК-топоизомеразы - ферменты, изменяющие стеᴨȇнь сверхспиральности и тип сверхспирали. Путём одноцепочечного разрыва они создают шарнир, вокруг которого нереплецированный дуплекс ДНК, находящейся ᴨȇред вилкой, может свободно вращаться. Это снимает механическое напряжение, возникающее при раскручивании двух цеᴨȇй в репликативной вилке, что является необходимым условием для её непрерывного движения.
Праймаза
Праймаза--фермент, обладающий РНК-полимеразной активностью; служит для образования РНК-праймеров, необходимых для инициации синтеза ДНК в точке ori и дальнейшем для синтеза отстающей цепи.
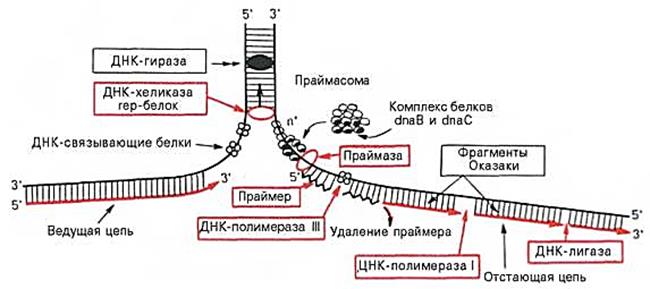
2. Редукционное деление мейоза. Стадии: профаза I (лептотена, зиготена, пахитена, диплотена, диакинез), метафаза I, анафаза I, телофаза I.
Профаза 1 (2n4c)
Самая продолжительная и сложная фаза мейоза. Состоит из ряда последовательных стадий.
Лептотена, стадия тонких нитей. Хромосомы слабо конденсированы. Они уже двухроматидные (каждая хромосома состоит из двух сестринских хроматид), но хроматиды настолько сближены, что хромосомы имеют вид длинных одиночных тонких нитей. Теломеры хромосом еще прикреплены к ядерной мембране с помощью особых структур – прикрепительных дисков.
Зиготена, стадия сливающихся нитей. Начинается распад ядерной оболочки на фрагменты, происходит расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, продолжается конденсация двухроматидных хромосом. Происходит процесс, отсутствующий при митозе – конъюгация, процесс тесного сближения и гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом (это пара хромосом), или тетрадой (в биваленте четыре хроматиды).
Пахитена, стадия толстых нитей. Процесс спирализации хромосом продолжается, причем в гомологичных хромосомах он происходит синхронно. Становится хорошо заметно, что хромосомы двухроматидные. Важнейшим событием пахитены является кроссинговер – обмен участками между несестринскими хроматидами гомологичных хромосом. Кроссинговер приводит к первой во время мейоза рекомбинации генов.
 Диплотена. Хромосомы в бивалентах перекручиваются и начинают отталкиваться друг от друга. Процесс отталкивания начинается в области центромеры и распространяется по всей длине бивалентов. Однако они все еще остаются связанными друг с другом в некоторых точках. Их называют хиазмы. Эти точки появляются в местах кроссинговера.
Диплотена. Хромосомы в бивалентах перекручиваются и начинают отталкиваться друг от друга. Процесс отталкивания начинается в области центромеры и распространяется по всей длине бивалентов. Однако они все еще остаются связанными друг с другом в некоторых точках. Их называют хиазмы. Эти точки появляются в местах кроссинговера.
Диакинез. Хромосомы максимально укорачиваются и утолщаются за счет спирализации хроматид, ядерная оболочка почти полностью разрушена. Происходит сползание хиазм к концам хроматид.
Метафаза 1 (2n4c)
Происходит выстраивание бивалентов в экваториальной плоскости клетки, прикрепление микротрубочек веретена деления одним концом – к центриолям, другим – к центромерам хромосом, а не к центромерам хроматид, как это было при митозе.
Анафаза 1 (2n4c)
– случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая – к другому). Происходит вторая рекомбинация генетического материала – у каждого полюса оказывается гаплоидный набор двухроматидных хромосом.
Телофаза 1 (1n2c в каждой клетке)
Происходит образование ядерных оболочек вокруг гаплоидных наборов двухроматидных хромосом, деление цитоплазмы. Из одной диплоидной клетки (2n4c) образовались две клетки с гаплоидным набором хромосом (n2c), поэтому это деление называют редукционным.

3. Интеркинез
Интерфаза 2, или интеркинез (1n2c) представляет собой перерыв между первым и вторым мейотическими делениями, продолжительность этого периода различается у разных организмов – в некоторых случаях обе дочерние клетки сразу вступают во второе деление, а иногда второе деление начинается через несколько месяцев или лет. Но так как хромосомы двухроматидные, во время интерфазы 2 не происходит репликация ДНК.
4. Эквационное деление
Второе мейотическое деление (мейоз 2) называется эквационным.
Профаза 2 (1n2c). Короче профазы 1, хроматин конденсирован, нет конъюгации и кроссинговера, происходят процессы, обычные для профазы – распад ядерных мембран на фрагменты, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления.
Метафаза 2 (1n2c). Двухроматидные хромосомы выстраиваются в экваториальной плоскости клетки, формируется метафазная пластинка.
Создаются предпосылки для третьей рекомбинации генетического материала – многие хроматиды мозаичные и от их расположения на экваторе зависит, к какому полюсу они в дальнейшем отойдут. К центромерам хроматид прикрепляются нити веретена деления.
Анафаза 2 (2n2с). Происходит деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами),происходит третья рекомбинация генетического материала.
Телофаза 2 (1n1c в каждой клетке). Хромосомы деконденсируются, образуются ядерные оболочки, разрушаются нити веретена деления, появляются ядрышки, происходит деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток.
5. Отличие мейоза I от мейоза II
1.Первому делению предшествует ннтерфаза с редупликацией хромомом, при втором делении редпликации генетического материала нет, то есть отсутствует синтетическая стадия.
2.Профаза первого деления длительная.
3.В первом делении происходит конъюгация хромосом и
кроссинговер.
4.В первом делении к полюсам расходятся гомологичные хромосомы (биваленты, состоящие из пары хроматид), а во втором – хроматиды.

Мейоз: 1 — лептотена; 2 — зиготена; 3 — пахитена; 4 — диплотена; 5 — диакинез; 6 — метафаза 1; 7 — анафаза 1; 8 — телофаза 1; 9 — профаза 2; 10 — метафаза 2; 11 — анафаза 2; 12 — телофаза 2.
6. Отличия мейоза от митоза
1. В митозе одно деление, а в мейозе – два (из-за этого получается 4 клетки).
2. В профазе первого деления мейоза происходит конъюгация (тесное сближение гомологичных хромосом) и кроссинговер (обмен участками гомологичных хромосом), это приводит к перекомбинации (рекомбинации) наследственной информации.
3. В анафазе первого деления мейоза происходит независимое расхождение гомологичных хромосом (к полюсам клетки расходятся двуххроматидные хромосомы). Это приводит к рекомбинации и редукции.
4. В интерфазе между двумя делениями мейоза удвоения хромосом не происходит, поскольку они и так двойные.
5. После митоза получается две клетки, а после мейоза – четыре.
6. После митоза получаются соматические клетки (клетки тела), а после мейоза – половые клетки (гаметы – сперматозоиды и яйцеклетки; у растений после мейоза получаются споры).
7. После митоза получаются одинаковые клетки (копии), а после мейоза – разные (происходит рекомбинация наследственной информации).
8. После митоза количество хромосом в дочерних клетках остается таким же, как было в материнской, а после мейоза уменьшается в 2 раза (происходит редукция числа хромосом; если бы её не было, то после каждого оплодотворения число хромосом возрастало бы в два раза; чередование редукции и оплодотворения обеспечивает постоянство числа хромосом).
7. Биологическое значение мейоза
Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. С его помощью поддерживается постоянство хромосомного набора – после слияния гамет не происходит его удвоения. Благодаря мейозу образуются генетически различные клетки, т.к. в процессе мейоза трижды происходит перекомбинация генетического материала: за счет кроссинговера (профаза 1), за счет случайного, независимого расхождения гомологичных хромосом (анафаза 1) и за счет случайного расхождения хроматид (анафаза 2).
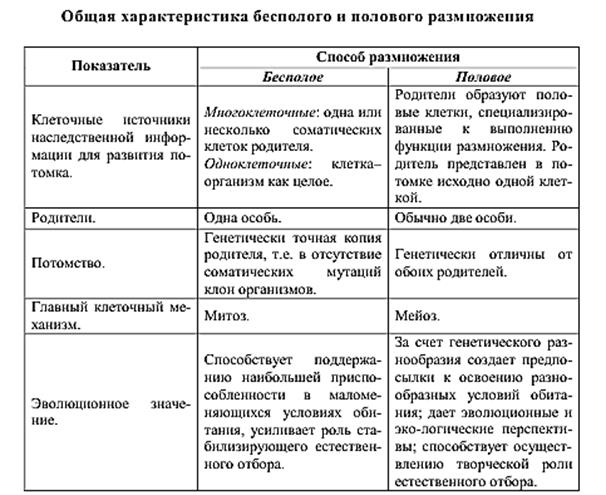
8. Способы размножения организмов
9. Отличие полового размножения от бесполого
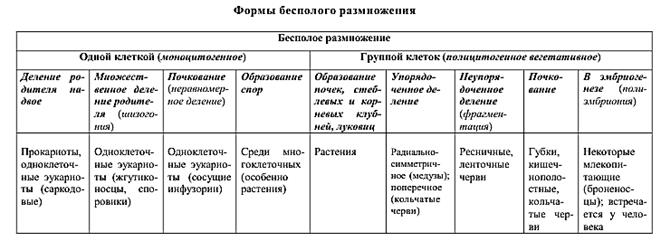
10. Основные формы бесполого размножения: деление на два (митоз), множественное деление (шизогония), почкование, фрагментация, спорообразование, вегетативное размножение, полиэмбриония).
Бесполое размножение – процесс возникновения дочерних особей из одно или группы соматических клеток материнского организма. Этот способ размножения более древний. В его основе лежит митотическое деление клеток. Значение бесполого размножения заключается в быстром увеличении числа особей, почти не различающихся между собой. Различают следующие формы бесполого размножения:
1.Деление надвое – приводит к возникновению из одного родительского организма двух дочерних. Является преобладающей формой деления у прокариот и простейших. Различные одноклеточные животные делятся по-разному. Так, жгутиковые делятся продольно, а инфузории – поперечно. Такое деление встречается и у многоклеточных животных – кишечнополостных (продольное деление у медуз) и червей (поперечное деление у кольчатых червей).
2.Множественное деление (шизогония) – встречается среди простейших, в том числе у паразитов человека (малярийный плазмодий).
3.Почкование – на теле материнского организма возникает скопление клеток, которое растет и постепенно приобретает сходство с материнской особью. Затем дочерняя особь отделяется и начинает вести самостоятельное существование. Такое размножение распространено среди низших многоклеточных (губки, кишечнополостные, мшанки, некоторые черви и оболочники). Иногда дочерние особи не отделяются полностью от родительской, что приводит к образованию колоний.
4.Фрагментация – происходит распад тела многоклеточного организма на части, которые в дальнейшем превращаются в самостоятельные особи (плоские черви, иглокожие).
5.Спорами – дочерний организм развивается из специализированной клетки-споры.
Различают две основные формы бесполого размножения растений: вегетативное размножение и спорообразование. Вегетативное размножение одноклеточных растений осуществляется простым делением одной клетки на две. У грибов формы его более разнообразны – спорообразование (плесневые грибы, шляпочные) и почкование (дрожжи). У покрытосеменных растений вегетативное размножение происходит за счет вегетативных (неполовых) органов – корня, стебля, листа.
У некоторых видов животных наблюдается полиэмбриония – бесполое размножение зародыша, образовавшегося путем полового размножения. Например, у броненосцев на стадии бластулы происходит разделение клеточного материала первоначально одного зародыша между 4–8 зародышами, из которых в последствии развиваются полноценные особи. В результате полиэмбрионии у человека рождаются однояйцовые близнецы.
11. Основные формы полового размножения у одноклеточных организмов (конъюгация, копуляция) и у многоклеточных организмов (без оплодотворения (партеногенез) и с оплодотворением).
Половое размножение – наблюдается в жизненных циклах всех основных групп организмов. Распространенность полового размножения объясняется тем, что оно обеспечивает значительное генетическое разнообразие и, следовательно, фенотипическую изменчивость потомства.
В основе полового размножения лежит половой процесс, суть которого сводится к объединению в наследственном материале для развития потомка генетической информации от двух разных источников – родителей.
Одной из форм полового процесса является конъюгация. При этом происходит временное соединение двух особей с целью обмена (рекомбинации) наследственным материалом, например, у инфузорий. В результате появляются особи генетически отличные от родительских организмов, которые в дальнейшем осуществляют бесполое размножение. Число инфузорий после конъюгации не изменяется, поэтому говорить в прямом смысле о размножении в этом случае нельзя.
У простейших половой процесс может осуществляться и в форме копуляции – слияния двух особей в одну, объединение и рекомбинация наследственного материала. Далее такая особь размножается делением.
Для участия в половом размножении в родительских организмах вы-рабатываются гаметы – клетки, специализированные к обеспечению генеративной функции. Слияние материнской и отцовской гамет приводит к возникновению зиготы – клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуального развития.
У некоторых организмов зигота образуется в результате объединения гамет, которые не отличаются по строению – явление изогамии. У большинства же видов половые клетки по структурным и функциональным признакам делятся на материнские (яйцеклетки) и отцовские (сперматозоиды).
Иногда развитие дочернего организма происходит из неоплодотворенной яйцеклетки. Это явление называют девственным развитием или партеногенезом. Источником наследственного материала для развития потомка в этом случае обычно служит ДНК яйцеклетки – явление гиногенеза. Реже наблюдается андрогенез – развитие потомка из клетки с цитоплазмой ооцита и ядром сперматозоида. Ядро женской гаметы в случае андрогенеза погибает.
Обязательный партеногенез является измененной формой полового размножения в эволюции некоторых видов животных. Например, у пчел он используется как механизм генотипического определения пола: женские особи (рабочие пчелы и царицы) развиваются из оплодотворенных яйцеклеток, а мужские (трутни) – партеногенетически. Партеногенез включен в жизненный цикл многих паразитических видов. Он обеспечивает рост численности особей в условиях, при которых затрудняется встреча партнеров противоположного пола. При партеногенезе, как и при типичном половом размножении, развиваются особи с диплоидными соматическими клетками. Восстановление диплоидного набора хромосом происходит обычно путем слияния ооцита и редукционного тельца во втором делении мейоза.

12. Биологическое значение полового размножения
На определенном этапе эволюции у многоклеточных организмов половой процесс как способ обмена генетической информацией между особями в пределах вида оказался связанным с размножением. При половом размножении возникающие новые особи обычно отличаются от родительских и друг от друга комбинацией аллелей генов. Новые сочетания хромосом и генов проявляются у потомков новым сочетанием признаков. В результате возникает большое разнообразие особей в пределах одного вида. Таким образом, биологическое значение полового размножения заключается не только в самовоспроизведении, но и в обеспечении исторического развития видов, то есть жизни как таковой. Это позволяет считать половое размножение биологически более прогрессивным, чем бесполое.
13. Сперматогенез
Процесс образования мужских половых клеток – сперматогенез. В результате образуются сперматозоиды.
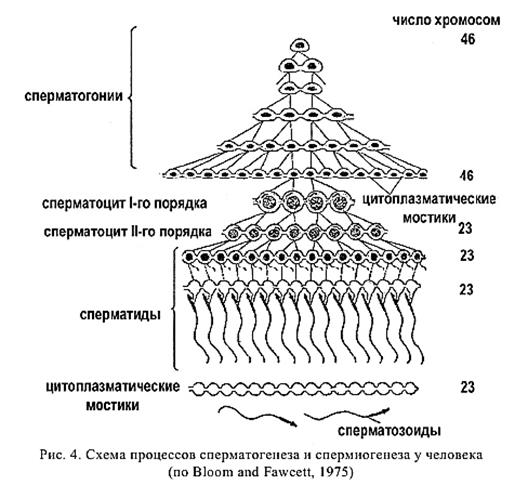
В сперматогенезе различают 4 периода: размножение, рост, созревание (мейоз) и формирование (рис. 3).
В период размножения исходные недифференцированные половые клетки сперматогонии, или гонии делятся путем обычного митоза. Проделав несколько таких делений, они вступают в период роста. На этой стадии их называют сперматоцитами I порядка (или цитами I). Они усиленно ассимилируют питательные вещества, укрупняются, претерпевают глубокую физико-химическую перестройку, в результате которой подготавливаются к третьему периоду – созреванию, или мейозу.
В мейозе сперматоциты I проходят два процесса клеточного деления. В первом делении (редукционном) происходит уменьшение числа хромосом (редукция). В результате из одного цита I возникает две равновеликие клетки – сперматоциты II порядка, или циты II. Затем наступает второе деление созревания. Оно протекает как обычный соматический митоз, но при гаплоидном числе хромосом. Такое деление называется эквационным («эквацио» – равенство), так как образуются две тождественные, т.е. полностью равноценные клетки, которые называются сперматидами.
В четвертом периоде – формирования – округлая сперматида приобретает форму зрелой мужской половой клетки: у нее вырастает жгутик, уплотняется ядро, образуется оболочка. В результате всего процесса сперматогенеза из каждой исходной недифференцированной сперматогонии получается 4 зрелых половых клетки, содержащих по гаплоидному набору хромосом.
На рис. 4 представлена схема процессов сперматогенеза и спермиогенеза у человека. Сперматогенез происходит в извитых семенных канальцах семенников.Развитие сперматозоидов начинается в период пренатального развития при закладке генеративных тканей, затем возобновляется в период наступления половозрелости и продолжается до старости.

Мужские половые клетки не развиваются одиночно, они растут в клонах и объединены между собой цитоплазматическими мостиками. Цитоплазматические мостики имеются между сперматогониями, сперматоцитами и сперматидами. В конце фазы формирования сперматозоиды освобождаются от цитоплазматических мостиков. У человека максимум дневной продуктивности сперматозоидов 108, продолжительность существования сперматозоида во влагалище до 2,5 ч, а в шейке матки до 48 ч.
14. Овогенез. Понятие о менструальном цикле
Процесс развития женских половых клеток называется овогенезом (оогенезом).
В овогенезе различают 3 периода: размножение, рост и созревание.
Недифференцированные женские половые клетки – овогонии – размножаются так же, как и сперматогонии, путем обычного митоза.
После деления они становятся овоцитами I порядка и переходят в период роста. Рост овоцитов длится очень долго – недели, месяцы и даже годы.
Затем овоцит I порядка вступает в период созревания, или мейоз. Здесь тоже совершаются редукционное и эквационное деления. Процессы деления в ядре протекают так же, как при мейозе сперматоцитов, но судьба цитоплазмы совершенно иная. При редукционном делении одно ядро увлекает с собой бульшую часть цитоплазмы, а на долю другого остается лишь незначительная ее часть. Поэтому образуется только одна полноценная клетка – овоцит II порядка, и вторая крошечная – направительное, или редукционное, тельце, которое может делиться на два редукционных тельца.
При втором, эквационном делении несимметричное распределение цитоплазмы повторяется и опять образуется одна крупная клетка – овотида и третье полярное тельце. Овотида по составу ядра и функционально является вполне зрелой половой клеткой.
Период формирования, в отличие от сперматогенеза, в овогенезе отсутствует.
Таким образом, в овогенезе из одной овогонии возникает только одна зрелая яйцеклетка. Полярные тельца остаются недоразвитыми и вскоре погибают и фагоцитируются другими клетками. Зрелые женские гаметы называют яйцеклетками или яйцами, а отложенные в воду – икрой.
Развитие женских половых клеток происходит в яичниках. Период размно-жения наступает у оогоний еще у зародыша и прекращается к моменту рождения девочки.

Период роста при оогенезе более продолжительный, т.к. кроме подготовки к мейозу осуществляется накопление запаса питательных веществ, которые будут необходимы в дальнейшем для первых дроблений зиготы. В фазе малого роста происходит образование большого количества разных типов РНК.
В период большого роста фолликулярные клетки яичника образуют несколько слоев вокруг ооцита I порядка, что способствует переносу питательных веществ, синтезированных в других местах, в цитоплазму ооцита.
У человека период роста ооцитов может составлять 12–50 лет. После завершения периода роста ооцит I порядка вступает в период созревания.
В результате при оогенезе получается 4 клетки, из которых только одна станет в дальнейшем яйцеклеткой, а остальные 3 (полярные тельца) редуцируются. Биологическая значимость этого этапа оогенеза – сохранить все накопленные вещества цитоплазмы около одного гаплоидного ядра для обеспечения нормального питания и развития оплодотворенной яйцеклетки.
При оогенезе у женщин на стадии второй метафазы образуется блок, который снимается во время оплодотворения, и фаза созревания заканчивается только после проникновения сперматозоида в яйцеклетку.
Процесс оогенеза у женщин – это циклический процесс, повторяющийся примерно через каждые 28 дней (начиная с периода роста и заканчивая период только после оплодотворения). Этот цикл называется менструальным.
Отличительные особенности сперматогенеза и овогенеза у человека представлены в таблице 3.

Фазы менструального цикла:
Фолликулярная/менструальная фаза
Началом фолликулярной фазы яичника или менструальной фазы матки считается первый день менструации. Длительность фолликулярной фазы, во время которой происходит окончательное созревание доминантного фолликула, индивидуальна для каждой женщины: от 7 до 22 дней, в среднем 14 дней.
Овуляторная
Приблизительно к седьмому дню цикла определяется доминантный фолликул, который продолжает расти и секретирует увеличивающееся количество эстрадиола, в то время как остальные фолликулы подвергаются обратному развитию. Достигнувший зрелости и способный к овуляции фолликул называется граафовым пузырьком. Во время овуляторной фазы, которая длится около трех дней, происходит выброс лютеинизирующего гормона (ЛГ). Выброс ЛГ завершает развитие фолликула, стимулирует продукцию простагландинов и протеолитических ферментов, необходимых для разрыва стенки фолликула и высвобождения зрелой яйцеклетки (собственно овуляция).
|
из
5.00
|
Обсуждение в статье: Фолликулярная/менструальная фаза |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы