 |
Главная |


Возрастная изменчивость фитоценозов
|
из
5.00
|
Возрастная измененчивость фитоценозов, по В. Н. Сукачеву, это "изменение фитоценозов в связи с онтогенезом эдификаторов". Как уже было сказано, отдельные особи, входящие в состав популяции вида, в связи с различиями в возрасте и жизненном состоянии сильно отличаются друг от друга по массе подземных и надземных органов и по интенсивности влияния на среду и другие растения. Следовательно, роль вида в фитоценозе определяется не только его численностью, экологическими особенностями, но и возрастным составом его популяции. А так как возрастной состав год от года не остается одинаковым, изменяются и особенности фитоценоза. Не следует забывать и о том, что внешними условиями существования вида в фитоценозе определяется и длительность прохождения им отдельных возрастных стадий, и урожайность семян, способность семян прорастать и т.д.
Возрастные изменения фитоценозов удобнее всего рассматривать на примере лесных фитоценозов. Дело в том, что в лесах изменения структуры и состава видов в фитоценозе происходят при сохранении лесообразующего значения доминирующего вида деревьев. Эти изменения протекают "внутри" лесного фитоценоза и охватывают промежуток времени, как минимум, равный продолжительности жизни одного поколения доминирующей древесной породы (обычно от 40 до 300 лет). Эти смены определяются изменениями режимов различных экологических факторов, которые, однако, не выходят за пределы определенной нормы, характерной для данного типа леса. При этом происходит закономерное повторение определенных морфологически однородных стадий развития лесного фитоценоза.
Примером онтогенетических изменений может служить развитие лесного сообщества от стадии молодняка до приспевающего и спелого леса. В результате подобного развития происходит не только изменение возраста деревьев-эдификаторов, но и соответствующие изменения экологических условий фитоценоза, а также изменяется соотношение различных компонентов растительного покрова. При этом не стоит забывать о том, что данные изменения не выходят за рамки одного фитоценоза, так, например, ельник-кисличник и возрасте 30-40 лет, и возрасте 150-180 лет остается все тем же ельником-кисличником. На лугах или в степях онтогенетические изменения возрастного состава доминирующих растений, также как и в лесах, будут сопровождаться соответствующими сменами сопутствующих видов.
С онтогенетическими изменениями очень тесно связаны процессы возобновления внутри сообществ, например, замена отмирающих особей другими. Так, одной из характерных особенностей дождевого тропического леса является непрерывное из года в год изменение в расположении отдельных видов деревьев в древостое. Дело в том, что в подобном растительном сообществе вероятность вырастания дерева того же вида на месте погибшего крайне низка, так как число лесообразующих видов деревьев очень велико. Это приводит к тому, что через 20-25 лет расположение деревьев в фитоценозе будет в значительной степени иным по сравнению с исходным. Естественно, что вслед за сменой вида дерева меняется и связанная с его экологическими особенностями микрогруппировка.
Все перечисленные выше изменения растительности происходят в пределах одного фитоценоза. В то же время эти изменения могут выйти за его пределы. Так, разногодичные изменения при увеличении амплитуды колебаний условий существования могут перейти в смены одного сообщества другим. Смены, вызванные возобновлением того или иного вида, при гибели большей части генеративных особей также способны привести к смене фитоценозов. В самом общем случае эти изменения называются сукцессионными процессами.
Сукцессии
В природе отсутствуют фитоценозы, которые бы существовали вечно. Рано или поздно один фитоценоз либо сменяется другим, либо уничтожается под воздействием каких-либо внешних причин, включая деятельность человека. Подобная ограниченность существования фитоценозов во времени, их сменяемость является одним из важнейших свойств растительного покрова. Это возможно в силу того, что фитоценозы являются открытыми системами, которые непрерывно испытывают воздействие меняющихся по своей интенсивности внешних факторов (космических, климатических, гидрологических, геоморфологических, антропических и т.д.). Кроме того, фитоценозы являются системами, открытыми для внедрения новых видов растений и других организмов.
Все перечисленные выше причины и вызывают смены одних фитоценозов другими. Если нет изменений в воздействии внутренних и внешних факторов, то эти изменения необратимы и происходят в определенном направлении. Такой тип изменений называется сукцессиями.
Сукцессии – постепенные направленные, как правило,необратимые изменения растительности, которые могут быть вызваны как внутренними, так и внешними причинами, и при которых изменяется видовой состав или количественное соотношение видов, то есть структура фитоценозов. Именно направленность и масштаб изменений – основное, что отличает сукцессии от флуктуаций.
Сукцессии включают в себя очень обширный и разнородный спектр процессов, происходящих в живой природе, и поэтому они нуждаются в какой-то классификации. Сукцессии можно классифицировать по достаточно большому числу критериев, например:
по масштабу времени – на:
· быстрые, протекающие в масштабе десятилетий,
· средние,длящиеся столетиями,
· медленные, продолжающиеся тысячелетиями,
· очень медленные(десятки тысяч лет);
по обратимости – на:
· обратимые
· необратимые;
по степени постоянства процесса – на:
· постоянные
· прерывающиеся;
по происхождению – на:
· первичные, то есть те, которые начинаются на субстратах, где никогда не было растительности,
· вторичные,происходящие на месте уже сформировавшихся фитоценозов после их нарушения или уничтожения;
по характеру изменения структуры и видового состава – на:
· прогрессивные(в результате сукцессии увеличивается видовое богатство, повышается продуктивность)
· регрессивные(в результате сукцессии уменьшается видовое богатство и снижается продуктивность)
Следует отметить, что между видовым богатством фитоценоза и его продуктивностью нет жесткой положительной зависимости и поэтому иногда при обеднении видового богатства фитоценоза его продуктивность может, наоборот, увеличиваться. В таких случаях по тенденциям изменения видового богатства и продуктивности отдельно различают прогрессивные и регрессивные сукцессии.
по антропогенности – на
· антропогенные (вызванные влиянием человека)
· природные (происходящие по естественным причинам)
по причинам – на
· автогенные –изменения фитоценозов вызываются внутренними по отношению к фитоценозу причинами (взаимоотношения растений, отношения растений к условиям среды, и т.д.)
· аллогенные – изменения фитоценозов вызываются внешними по отношению к фитоценозу причинами.
Кроме этого, по предложению В. Н. Сукачева различают по два типа автогенных и аллогенных сукцессий.
Автогенные сукцессии разделяются на сингенез и эндоэкогенез. При сингенезе изменения происходят под влиянием взаимоотношений между растениями. Этот быстрый процесс можно наблюдать главным образом на богатом субстрате. В данном случае под влиянием наличия или отсутствия банка семян в почве, способа доставки семян к месту сукцессии, длительности жизни за несколько лет происходит формирование фитоценоза, состоящего главным образом из однолетников-эксплерентов. В дальнейшем растения-однолетники постепенно заменяются многолетниками и им на смену приходят виды, сочетающие в себе особенности эксплерентов, патиентов и виолентов. При эндоэкогенезе в результате жизнедеятельности растений изменяются условия среды. Примерами подобных сукцессий могут служить процессы, происходящие при формировании почвы на скалах, зарастании озер и т.д.
Оба типа сукцессий очень тесно связаны. При сингенезе также происходит изменение условий среды, но, по сравнению с ролью взаимоотношений растений, роль этих изменений незначительна, так как почва не успевает существенно и, тем более, необратимо измениться. При эндоэкогенезе взаимоотношения между растениями также определенным образом влияют на изменения в фитоценозе, но гораздо слабее, чем преобразование фитоценозом собственной среды обитания.
Аллогенные сукцессии разделяются на гологенез и гейтогенез. Гологенез –сукцессии, являющиеся следствием процессов развития ландшафтов, обусловленных как природными, так и антропогенными причинами (например, формирование речной поймы, строительство водохранилищ и т.д.). Гейтогенез – сукцессии, обусловленные внешними факторами, которые не связаны с общими тенденциями развития ландшафта и имеют, главным образом, антропогенную природу.
Автогенные сукцессии.
Автогенные сукцессии очень разнообразны, они могут быть первичными и вторичными, прогрессивными и регрессивными, быстрыми и медленными. Как правило, они необратимы и протекают постоянно. Несмотря на спонтанность этих сукцессий, их может вызвать также и человек, нарушающий растительность. При этом на месте нарушения начинаются восстановительные автогенные сукцессии.
Все разнообразие автогенных сукцессионных процессов можно свести к четырем принципиальным моделям сукцессии, которые были предложены Дж. Коннелом и П. Слэйтьером (Connel, Slatyer, 1977) и Д. Боткиным (Botkin, 1981).
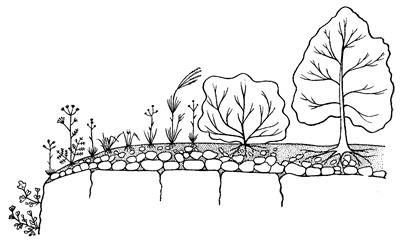 Модель благоприятствования. В ходе сукцессии, протекающей по этой модели, смена видов связана с постепенным улучшением условий среды, и поэтому она протекает как прогрессивная. Такую сукцессию при зарастании скал изучал один из крупнейших американских фитоценологов, основоположник учения о сукцессиях Ф. Клементс. Для процесса зарастания скал он выделял стадии накипных лишайников, кустистых лишайников и мхов, трав, кустарников, деревьев. Растения и связанные с ними гетеротрофные организмы в ходе сукцессии этого типа постепенно преобразуют горную породу и создают почву. По мере улучшения условий среды возрастает продуктивность фитоценоза, усложняется его структура, появляется ярусность и повышается видовое богатство (рис. 29).
Модель благоприятствования. В ходе сукцессии, протекающей по этой модели, смена видов связана с постепенным улучшением условий среды, и поэтому она протекает как прогрессивная. Такую сукцессию при зарастании скал изучал один из крупнейших американских фитоценологов, основоположник учения о сукцессиях Ф. Клементс. Для процесса зарастания скал он выделял стадии накипных лишайников, кустистых лишайников и мхов, трав, кустарников, деревьев. Растения и связанные с ними гетеротрофные организмы в ходе сукцессии этого типа постепенно преобразуют горную породу и создают почву. По мере улучшения условий среды возрастает продуктивность фитоценоза, усложняется его структура, появляется ярусность и повышается видовое богатство (рис. 29).
| Рис. 29. Схема сукцессии на известняковых скалах в районе Праги, южная экспозиция (по Клике). |
Этой же модели соответствует естественный процесс зарастания отвалов пустой породы на месте горных выработок, если порода является благоприятным субстратом для жизни растений. Если же она непригодна для растений, то человек может помочь началу сукцессии, покрывая поверхность породы слоем почвы или торфа. Ускорить сукцессию, протекающую по модели благоприятствования, можно высевом травосмеси или внесением удобрений, что практиковали американские экологи на Аляске при восстановлении растительного покрова после нефтяных разработок.
Модель толерантности. Смена видов в ходе сукцессии, протекающей по этой модели, происходит в результате ухудшений условий среды. Подобные сукцессии в природе распространены, по-видимому, не менее часто, чем сукцессии, соответствующие предыдущей модели. Как правило, подобные сукцессии наблюдаются тогда, когда растения, заселяя местообитания с исходно благоприятными условиями среды, постепенно расходуют ресурсы, что приводит к усилению конкурентных взаимоотношений между ними. В результате видовой состав меняется в направлении усиления роли видов с выраженным свойством патиентности.
Примером проявления модели толерантности может служить сукцессия, происходящая на вырубке в том случае, если она начинает использоваться в качестве сенокоса или пастбища. При этом начинают формироваться сенокосные или пастбищные травостои, типичные для данной зоны. Так, к примеру, в лесной зоне на смену крупнотравью, состоящему из иван-чая, таволги вязолистной и др. и развивающемуся вследствие минерализации лесной подстилки и корней вырубленных деревьев, приходят злаково-разнотравные фитоценозы с участием вейников, пырея ползучего, овсяницы луговой, ежи сборной. По мере того, как с урожаем из почвы выносятся элементы минерального питания, продуктивность фитоценоза постепенно снижается и крупные злаки сменяются мелкими – полевицей тонкой, трясункой средней, душистым колоском.
Если же продолжается использование этого фитоценоза и дальше, то при продолжении сенокошения в нем постепенно начинает доминировать белоус торчащий; если же такой фитоценоз используется как пастбище, то формируются сообщества, состоящие главным образом из манжеток. Белоусники и манжетковые луга – это самые бедные луговые сообщества, практически непригодные ни для какого дальнейшего использования человеком. Именно поэтому описанная сукцессия, протекающая по модели толерантности, крайне нежелательна. Применяя удобрения и правильно используя вырубки, можно воспрепятствовать этому и задержать сукцессию на стадии ценных в кормовом отношении и продуктивных злаков.
Еще одним примером сукцессии, протекающей по модели толерантности, может служить процесс самовосстановления елового леса на месте вырубки. Как правило, в подобных условиях вначале формируются мелколиственные леса, состоящие из ольхи, березы или ивы. Затем под их пологом поселяется более устойчивая к затенению и, соответственно, с более выраженным свойством патиентности ель, которая постепенно вытесняет мелколиственные породы (рис. 30). Под пологом ели развиваются еще большие патиенты – мхи и теневыносливые кустарнички и травы.
Модель ингибирования. Эта модель соответствует регрессивным сукцессиям, когда процесс приостанавливается в результате появления в фитоценозе видов, создающих условия, неприемлемые для 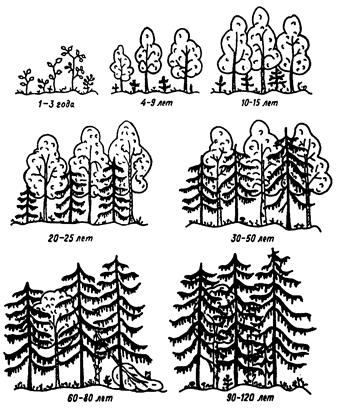 внедрения в фитоценоз новых видов. Сукцессии, протекающие по подобной модели, описаны, например, в Англии на гарях, где развивается густой покров кукушкина льна. При сильном развитии мохового покрова семена деревьев не могут достигнуть поверхности почвы и прорасти, поэтому такие фитоценозы десятки лет могут сохраняться в виде зеленомошной гари.
внедрения в фитоценоз новых видов. Сукцессии, протекающие по подобной модели, описаны, например, в Англии на гарях, где развивается густой покров кукушкина льна. При сильном развитии мохового покрова семена деревьев не могут достигнуть поверхности почвы и прорасти, поэтому такие фитоценозы десятки лет могут сохраняться в виде зеленомошной гари.
| Рис. 30. Схема восстановления елового леса на вырубке (Новиков, 1979). |
Аналогичный процесс происходит в пустынях Средней Азии, где при отсутствии выпаса поверхность почвы покрывается плотной коркой из мхов и водорослей (ее называют карахарсанг). В результате нарушается семенное возобновление пустынных деревьев и кустарников и происходит деградация всей экосистемы.
Характерно, что и на гари, и в пустыне сукцессия по модели ингибирования обусловлена тем, что из экосистем исключаются животные. Если моховую гарь посещают олени, то они копытами нарушают моховой покров и создают регенерационные ниши для возобновления деревьев. Подобным же образом пасущиеся в пустыне овцы и верблюды создают регенерационные ниши для поселения растений (Миркин и др., 2002).
Модель нейтральности. Этой модели соответствуют сукцессии, при которых изменения фитоценозов протекают как популяционный процесс, при котором происходит смена популяций видов с разными жизненными циклами и разными эколого-фитоценотическими типами стратегий. Роль взаимодействия популяций при данном типе сукцессий незначительна и предшественники практически не влияют на внедрение новых видов. Такие сукцессии крайне редки и чаще эта модель распространяется только на формирование видового состава, а количественные соотношения между видами обусловливаются ослабленным проявлением эффектов благоприятствования, ингибирования или толерантности.
Сукцессии, протекающие только по одной вышеизложенной модели, в природе, по-видимому, встречаются не так часто. В большинстве же случаев разные периоды сукцессионного процесса протекают в соответствии с разными моделями. При этом сукцессия, как правило, начинается с модели благоприятствования или нейтральности и заканчивается моделью толерантности. Это связано с тем, что на самых ранних стадиях автогенных сукцессий конкуренция между растениями чаще всего ослаблена. Но в дальнейшем даже тогда, когда осуществляется сильная биотическая трансформация местообитания и улучшение условий произрастания растений, происходит обострение конкурентных взаимоотношений между видами.
| Рис. 31. Схема строения болота, образовавшегося зарастанием озера (по Сукачеву): 1 – пресноводный мергель, 2 – сапропелит, 3 – сапропелевый торф, 4 – тростниковый торф, 5 – камышовый торф, 6 – хвощевый торф, 7 – осоковый торф, 8 – осоково-ивовый торф, 9 – лесной торф, 10 – гипновый торф, 11 – шейхцериево-сфагновый торф, 12 – сфагновый торф с пнями сосны, 13 – отложения мочажин на сфагновом торфе. |
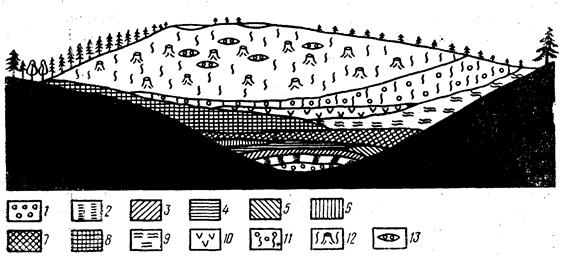 Примером сукцессии со сменой модели является процесс зарастания озера и последующего развития на его месте верхового болота (рис. 31). Вначале, когда чаша озера наполняется отмирающими растениями и сапропелем, в результате чего повышается уровень дна, условия для роста растений могут улучшаться и на смену плавающим макрофитам (рдестам, телорезу, ряскам, многокореннику) приходят высокие полупогруженные растения – тростник, рогоз, камыш озерный, хвощ болотный. Постепенно озеро окончательно зарастает и превращается в так называемое низинное, или эвтрофное (богатое) болото: на нем начинается процесс накопления торфа и поселяются осоки и ольха черная. В дальнейшем, в зависимости от особенностей местности, дальнейшее развитие заросших водоемов приводит или к возникновению лесных фитоценозов, или к образованию верхового болота, лишенного связи с грунтовыми водами, имеющего атмосферное питание и характеризующегося господством сфагновых мхов. Так при формировании верхового болота на смену растениям, требовательным к условиям питания, приходят виды, приспособленные к обитанию в условиях ограниченных ресурсов минеральных элементов: береза пушистая, вахта трехлистная, белокрыльник болотный. Это связано с тем, что год от года мощность слоя торфа увеличивается и растения, корни которых уже не достигают почвы, переходят на питание за счет торфа. Но, так как минерализация торфа на болоте происходит медленно и не до конца, каждому новому поколению растений достается все меньше питательных элементов. При дальнейшем нарастании торфяного слоя условия питания еще больше ухудшаются, и все болото покрывается сфагновым мхом. Низинное болото превращается в верховое. На нем появляется толерантная форма сосны, а также клюква, багульник болотный, болотный мирт, подбел и др.
Примером сукцессии со сменой модели является процесс зарастания озера и последующего развития на его месте верхового болота (рис. 31). Вначале, когда чаша озера наполняется отмирающими растениями и сапропелем, в результате чего повышается уровень дна, условия для роста растений могут улучшаться и на смену плавающим макрофитам (рдестам, телорезу, ряскам, многокореннику) приходят высокие полупогруженные растения – тростник, рогоз, камыш озерный, хвощ болотный. Постепенно озеро окончательно зарастает и превращается в так называемое низинное, или эвтрофное (богатое) болото: на нем начинается процесс накопления торфа и поселяются осоки и ольха черная. В дальнейшем, в зависимости от особенностей местности, дальнейшее развитие заросших водоемов приводит или к возникновению лесных фитоценозов, или к образованию верхового болота, лишенного связи с грунтовыми водами, имеющего атмосферное питание и характеризующегося господством сфагновых мхов. Так при формировании верхового болота на смену растениям, требовательным к условиям питания, приходят виды, приспособленные к обитанию в условиях ограниченных ресурсов минеральных элементов: береза пушистая, вахта трехлистная, белокрыльник болотный. Это связано с тем, что год от года мощность слоя торфа увеличивается и растения, корни которых уже не достигают почвы, переходят на питание за счет торфа. Но, так как минерализация торфа на болоте происходит медленно и не до конца, каждому новому поколению растений достается все меньше питательных элементов. При дальнейшем нарастании торфяного слоя условия питания еще больше ухудшаются, и все болото покрывается сфагновым мхом. Низинное болото превращается в верховое. На нем появляется толерантная форма сосны, а также клюква, багульник болотный, болотный мирт, подбел и др.
Аллогенные сукцессии
Аллогенные сукцессии обусловлены влиянием внешнего фактора, который действует на фитоценоз. При этом чаще всего по мере ухудшения условий среды происходит смена видов в направлении их более высокой патиентности и приспособленности к фактору, вызвавшему сукцессию, то есть, такие сукцессии соответствуют модели толерантности. Реже аллогенные сукцессии протекают по модели благоприятствования, например, при удобрении лугов или эвтрофикации (повышении содержания в воде питательных элементов) водоемов. Но в этом случае уменьшается видовое богатство фитоценозов. В его отсутствие аллогенная сукцессия сменяется автогенной восстановительной.
При гологенезе, как уже говорилось, вследствие обусловленных различными причинами процессов развития ландшафтов сопряженно изменяется растительность целого ландшафта или его существенной части. Такие сукцессии могут быть природными и антропогенными.
| Рис. 32. Зоны речной поймы средней полосы европейской части России: П – пионерных сообществ, И – ивы, Т – тополя, В – вяза, Л – липы, О – черной ольхи; А-Б - поперечный профиль (Миркин и др., 2002). |
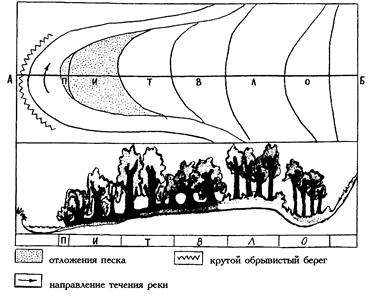 Как пример природной гологенетической сукцессии можно рассматривать процессы изменения растительности, происходящие при развитии речной долины (рис. 32). С одной стороны, текущая в реке вода постоянно размывает и углубляет ее русло, с другой стороны, поверхность поймы повышается за счет отложения взвешенных частиц, переносимых водой во время паводка (так называемого наилка). Кроме того, при меандрировании с одной стороны русла реки постоянно разрушается часть высокого берега, а с противоположной – наращивается песчаный пляж. В результате вышеописанных процессов на вновь образующихся намываемых субстратах происходит смена растительности по ряду: пионерные группировки – ивняк – осинник – вязовый лес – липово-дубовый лес. Кроме того, к террасе будет примыкать притеррасная пойма – область выклинивания грунтовых вод, где формируются низинные болота из Alnus glutinosa.
Как пример природной гологенетической сукцессии можно рассматривать процессы изменения растительности, происходящие при развитии речной долины (рис. 32). С одной стороны, текущая в реке вода постоянно размывает и углубляет ее русло, с другой стороны, поверхность поймы повышается за счет отложения взвешенных частиц, переносимых водой во время паводка (так называемого наилка). Кроме того, при меандрировании с одной стороны русла реки постоянно разрушается часть высокого берега, а с противоположной – наращивается песчаный пляж. В результате вышеописанных процессов на вновь образующихся намываемых субстратах происходит смена растительности по ряду: пионерные группировки – ивняк – осинник – вязовый лес – липово-дубовый лес. Кроме того, к террасе будет примыкать притеррасная пойма – область выклинивания грунтовых вод, где формируются низинные болота из Alnus glutinosa.
Вырубая леса в границах водосбора, человек может ускорять эту сукцессию. Вырубки, как правило, приводят к усиленной эрозии почвы, вследствие чего в пойму с весенними водами будет поступать большее количество смытой почвы. Это приведет к тому, что паводки станут более высокими, а процесс разрушения и созидания поймы активизируется. Облесением же водосбора человек может, напротив, снизить скорость процесса развития речной долины.
Как пример антропогенного гологенеза могут рассматриваться изменения растительности, происходящие в результате постройки крупных гидротехнических сооружений, например водохранилищ. Во первых, в результате затопления водами водохранилища наземных фитоценозов на их месте формируются водные сообщества. Во-вторых, выше плотины повышается уровень грунтовых вод и происходит подтопление почвы. Это приводит к тому, что существовавшие ранее на данной территории леса и травяная растительность сменяются фитоценозами, устойчивыми к высокому уровню грунтовых вод. Так, к примеру, вместо липово-дубовых, еловых или сосновых лесов формируются ольшаники, а вместо сухих лугов – осоковые болота. Ниже же плотины, наоборот, происходит понижение уровня грунтовых вод и, как следствие этого, иссушение поймы, а в южных районах – еще и засоление почв. В третьих, водохранилище изменяет климат, который становится более мягким и более ровным.
Оба рассмотренных примера показывают основную особенность гологенеза: под влиянием внешних факторов согласованно и закономерно изменяется совокупность фитоценозов на обширных территориях.
Гейтогенетические сукцессии – это изменения фитоценозов под влиянием действия внешних факторов, которые не связаны с общими тенденциями развития ландшафта. Как правило, основным фактором, вызывающим гейтогенетические сукцессии, является человек. В результате его деятельности происходят, главным образом, регрессивные сукцессии, называемые дигрессиями. Считается, что если в фитоценозе сохранилась хотя бы половина видов, то после прекращения действия фактора, вызвавшего дигрессивные изменения, возможно самовосстановление фитоценоза в результате процесса обратной трансформации, то есть вторичной автогенной сукцессии, которая называется восстановительной, или демутацией.
Рассмотрим некоторые примеры подобных сукцессий.
Сукцессии под влиянием выпаса (пастбищные дигрессии). Это самый распространенный вариант сукцессии. Перевыпас и, как следствие, происходящее при этом обеднение фитоценозов связаны с избыточным поголовьем скота. На сегодняшний день подобные сукцессии являются одним из самых тяжелых последствий хозяйственной деятельности человека.
Выпас действует на травяные фитоценозы двояко. С одной стороны, разные виды растений избирательно поедаются скотом, что в итоге приводит к изменению видового состава пастбищ. С другой стороны, чрезмерный выпас скота приводит к уплотнению почвы и изменению ее водного, воздушного и солевого режимов. В результате высокотравные и малоустойчивые к выпасу растения сменяются пастбищным низкотравьем: подорожником средним, клевером ползучим, одуванчиками, лапчаткой гусиной и др. При этом уменьшается видовое богатство фитоценоза, упрощается структура, снижается общее проективное покрытие.
Рассмотрим стадии пастбищной дигрессии.
1. Стадия недостаточного выпаса. Для этой стадии, когда пастбищная нагрузка на фитоценоз очень слаба или вообще отсутствует, характерно скопление большого количества отмерших надземных органов растений, что препятствует нормальному росту типичных луговых трав. Семенное размножение дерновинных злаков подавлено, места, освобождающиеся после отмирания старых дерновин, занимают главным образом корневищные злаки. Поскольку испарение с поверхности почвы из-за накопления опада уменьшается, происходит мезофитизация травостоя.
2. Стадия умеренного выпаса. Все дерновинные злаки хорошо развиты, много их молодых дерновин, что говорит об их успешном семенном размножении. Одно- и двулетние виды подавлены. В этой стадии луг дает максимальную продукцию.
3. Стадия чрезмерного выпаса. Начинают исчезать или очень сильно подавляться ценные в кормовом отношении виды растений. Вместо их в зависимости от условий окружающей среды начинают преобладать практически не поедаемые скотом виды: осоки и щучка на сырых лугах, крупные щавели на богатых почвах, либо полыни на сухих почвах.
4. Стадия полного сбоя, или выгона. Из-за сильного механического разрушения дерновины копытами скота начинают преобладать однолетники со стелющимися побегами – спорыши, мятлик однолетний, дивалы и др.
В результате пастбищной дигрессии продуктивность травостоев может снижаться в десятки раз. Воспрепятствовать столь нежелательным последствиям можно лишь внедрением пастбищеоборота, при котором нагрузка скота на пастбище нормируется и разным участкам периодически предоставляется отдых.
Рекреационные сукцессии – это сукцессии, протекающие в растительности под влиянием специфической деятельности человека в процессе отдыха: вытаптывание, выжигание, засорение фитоценозов и т.д. Основным фактором, влияющим на фитоценоз в данном случае, является вытаптывание. Поэтому рекреационная сукцессия будет отличаться от пастбищной, главным образом, тем, что в данном случае отбор будет идти на устойчивые к вытаптыванию виды без учета их кормовых качеств.
Я. П. Дидух и П. Г. Плюта (1994) предлагают выделять следующие уровни рекреационной нагрузки:
1. рекреация отсутствует;
2. хотя и имеется определенная рекреационная нагрузка, но тем не менее сообщество еще сохраняет целостность своей структуры;
3. происходит смена доминирующих видов;
4. интенсивная рекреационная нагрузка ведет к локальному исчезновению растительного покрова, что дает возможность появиться в значительном количестве однолетникам, чаще всего эфемерам;
5. сбои, отсутствие травяного покрова из-за сильного вытаптывания.
В качестве примера можно рассмотреть антропогенную дигрессию в лесных сообществах. В лесах на ранних стадиях рекреационной сукцессии видовое богатство фитоценозов вначале повышается, так как лесные травы замещаются более устойчивыми к вытаптыванию луговыми растениями. Однако при усилении вытаптывания видовое разнообразие падает, начинают преобладать растения розеточных форм, устойчивые к вытаптыванию. В дальнейшем сукцессия захватывает подлесок и даже древесный ярус, так как уплотнение почвы препятствует возобновлению деревьев, ухудшает их состояние и даже может привести к усыханию древостоя.
Сукцессии под влиянием радиации. Эти сукцессии изучали Г. Вудвелл и Р. Уиттекер на экспериментальном полигоне на острове Лонг-Айленд (США). Результаты их исследований показали, что с повышением радиационной нагрузки, задаваемой гамма-излучением, изменения растительности происходят в обратной последовательности к сукцессии по модели благоприятствования при зарастании скал. Вначале исчезают деревья, потом кустарники, затем травы, мхи и лишайники.
После катастрофы на Чернобыльской АЭС в окрестностях взорвавшегося реактора также произошло усыхание деревьев (эту зону назвали "бурым лесом"), но кустарники и травы сохранились.
Внешние воздействия, вызывающие аллогенные сукцессии, могут воздействовать на фитоценозы как прямо, так и косвенно. В последнем случае в роли посредников между абиотическими факторами и растениями могут выступать как автотрофные, так и гетеротрофные компоненты экосистемы. В качестве примера подобного опосредованного влияния можно рассмотреть влияние так называемых "кислых" дождей, когда с дождем в почву вносится довольно значительное количество связанного азота, на вересковые пустоши Шотландии. В результате проведенных исследований оказалось, что исчезновение из растительных сообществ вереска при воздействии «кислых» дождей вызывается, по меньшей мере, двумя факторами. Во-первых, оно связано с увеличением конкурентной способности нитрофилов (как правило, рудеральных заносных видов), во-вторых, с тем, что вереск, активно поглощая азот, начинает сильно повреждаться фитофагами.
Говоря о сукцессиях, нельзя не упомянуть о таком понятии, как "климакс" в растительности. Концепция климакса (устойчивого состояния, к которому "стремится" сообщество в ходе автогенной сукцессии) разрабатывалась американскими экологами еще в конце 19-го – начале 20-го столетия. Наиболее полно она была развита Ф. Клементсом, представления которого получили название моноклимакса в растительности.
Суть взглядов Ф. Клементса сводилась к следующему. В пределах одной климатической зоны все фитоценозы в ходе сукцессии должны конвергировать к одному климаксовому сообществу. Например, в умеренном климате при достижении определенной мощности торфяной залежи болота должны зарасти лесом, на скалах после образования почвы также должен вырасти лес и т.д. При этом в каждой климатической зоне в результате сукцессии все фитоценозы должны сойтись в один тип. Для тайги это будет еловый лес, для степной зоны – ковыльная степь, для тундр – сообщества мелких ив, осок и лишайников. Причем конечный результат данного процесса не будет зависеть от типа субстрата, на котором будут формироваться фитоценозы: на скалах, песках, глинах, торфянистых почвах в конечном итоге сформируются растительные сообщества одного состава, с одинаковой структурой и функциональными связями.
Ф. Клементс считал, что климаксовый фитоценоз формируется очень долго – тысячи и десятки тысяч лет, причем допускалась возможность различных отклонений от основного климакса, который назывался климатическим. Так, сообщества, возникающие в результате постоянного действия человека, препятствующего формированию климакса (например, выпас), назывались дисклимаксовыми. Если же формированию климакса мешали местные экологические факторы (например, паводки в пойме реки), то образованные в этом случае фитоценозы назывались субклимаксовыми и т.д. К концу жизни (Клементс публиковал свои работы в течение почти пятидесяти лет) он выделил около тридцати различных вариантов климакса. Климаксовые устойчивые фитоценозы Ф. Клементс противопоставлял серийным, т. е. сукцессионным. Смена серийных фитоценозов приводит к климаксу.
Представления Ф. Клементса о моноклимаксе стали предметом вполне справедливой критики. В результате была сформулирована концепция поликлимакса: в одной климатической зоне фитоценозы различных местообитаний изменяются в ходе сукцессии, но в один тип не конвергируют, а на песках, скалах, болотистых местообитаниях и т.д. формируются свои климаксовые сообщества.
В 50-х годах, когда растительный покров начал рассматриваться как непрерывное явление, Р. Уиттекером был предложен третий вариант концепции климакса – концепция климакс-континуума. Главной чертой, принципиально отличающей ее от других концепций, явилась констатация того, что между климаксовыми фитоценозами существуют переходы, и поэтому количество климаксов в поликлимаксе стремится к бесконечности. При этом в каждой точке под влиянием уникального сочетания факторов формируется свой индивидуальный климакс.
Р. Уиттекеру также принадлежит оригинальная классификация климаксов, в качестве основного критерия которой рассматривается длительность жизни доминантов серийных и климаксовых сообществ.
· Аклимакс. В данном случае время генерации доминантов меньше цикла изменений условий среды и флуктуации сообществ постоянны. Климаксовые и серийные сообщества в таком случае неразличимы. Примером аклимакса могут служить сообщества планктона.
· Циклоклимакс. Такой тип устойчивого состояния сообщества, в котором время генерации доминантов совпадает с годичными колебаниями условий среды. Как климаксовые сообщества, так и серийные в данном случае могут быть охарактеризованы главными доминантами-однолетниками. В качестве примера циклоклимаксовых сообществ могут рассматриваться некоторые сообщества пустынь, формируемые однолетними растениями.
· Катаклимакс. В данном случае генерация доминантов происходит в период между повторяющимися нарушениями среды, уничтожающими растительность. Это могут быть, к примеру, пожары, сели. Климаксовые и серийные сообщества в данном случае будут в большей или меньшей степени различаться.
· Суперклимакс, наблюдающийся, например, в тундре. В данных условиях формируется крайне низкая биомасса сообщества и более или менее стабильные популяции растений. При этом генерация доминантов длительна, а изменения среды незначительны. Все это приводит к тому, что серийные и климаксовые сообщества формируются из одних и тех же видов и различаются лишь своей структурой.
· Эуклимакс. По-видимому, наиболее широко распространенное устойчивое состояние фитоценоза, возникающее в условиях длительной генерации доминантов и более или менее непрерывно изменяющегося состава доминантов в ходе сукцессии. Данные фитоценозы характеризуются, как правило, высокой биомассой. Серийные сообщества при этом хорошо отличаются от климаксовых. Примером подобного процесса может служить классическая "клементсовская" сукцессия, которая приводит к развитию лесной растительности.
В современной фитоценологии концепция климакса играет важную роль, но климакс не абсолютизируется, а понимается как тенденция формирования фитоценозов зонального типа.
В заключение следует еще раз обратить внимание на то, что флуктуации, автогенные и аллогенные сукцессии не происходят независимо друг от друга, а представляют единый процесс изменения растительных сообществ. Это связано, в первую очередь, с тем, что лежащие в их основе процессы неотделимы друг от друга. Флуктуации, автогенные и аллогенные сукцессии есть составляющие непрерывного процесса изменения растительности. Различаются же они, главным образом, по степени обратимости изменений (флуктуации) и по преобладающим причинам, заключенным либо в самих сообществах (автогенные сукцессии), либо вне их (аллогенные сукцессии). Но всегда следует помнить, что, во-первых, при флуктуации не может быть полной обратимости, так как не прекращается автогенная сукцессия, в то же время в ходе аллогенных сукцессий всегда наблюдаются эндогенетические процессы.
|
из
5.00
|
Обсуждение в статье: Возрастная изменчивость фитоценозов |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы