 |
Главная |


Ферменты, участвующие в репликации ДНК
|
из
5.00
|
Процесс репликации осуществляется сложным ферментным комплексом, включающим 15-20 белков, которые условно разделяют на три группы.
1. Белки, подготавливающие родительскую ДНК к репликации
а) Узнающие (инициирующие) белки связываются со специфической последовательностью нуклеотидов в точке начала репликации и участвуют в присоединении к цепи ДНК всего комплекса ферментов репликации.
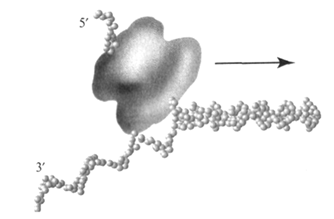 б) ДНК-хеликаза (от англ. helix – спираль) обеспечивает расплетание двойной спирали родительской ДНК в области репликативной вилки. Родительская ДНК разделяется на одноцепочечные участки, при этом затрачивается энергия гидролиза АТФ: 2 молекулы АТФ на разделение 1 пары нуклеотидов. Хеликаза, имеющая кольцевую структуру, однонаправленно перемещается по одной из цепей ДНК, расплетая перед собой двойную спираль, в результате чего возникает репликационная вилка (рис. 24). Вероятно, одновременно происходит вытеснение данного участка ДНК из связи с гистонами и другими хромосомными белками. Хеликазы часто функционируют в составе комплекса, осуществляющего перемещение репликативной вилки и репликацию расплетенных цепей.
б) ДНК-хеликаза (от англ. helix – спираль) обеспечивает расплетание двойной спирали родительской ДНК в области репликативной вилки. Родительская ДНК разделяется на одноцепочечные участки, при этом затрачивается энергия гидролиза АТФ: 2 молекулы АТФ на разделение 1 пары нуклеотидов. Хеликаза, имеющая кольцевую структуру, однонаправленно перемещается по одной из цепей ДНК, расплетая перед собой двойную спираль, в результате чего возникает репликационная вилка (рис. 24). Вероятно, одновременно происходит вытеснение данного участка ДНК из связи с гистонами и другими хромосомными белками. Хеликазы часто функционируют в составе комплекса, осуществляющего перемещение репликативной вилки и репликацию расплетенных цепей.
в) Белки, связывающиеся с одноцепочечной ДНК (SSB-белки – Single Strand Binding Proteins) обладают повышенным сродством к одноцепочечным участкам ДНК и стабилизируют их в таком состоянии. Роль SSB-белка в репликации, по-видимому, состоит в том, чтобы расправить ДНК и удалить возможные элементы вторичной структуры, которые могли бы образоваться в самокомплементарных участках ДНК. Связывание одноцепочечной ДНК с SSB-белком стимулирует ДНК-полимеразу и повышает точность ее работы.
г) Топоизомеразы. Каждая молекула ДНК зафиксирована в ядерном матриксе, поэтому она не может свободно вращаться при расплетании определенного участка. Это вызывает суперспирализацию (увеличивается количество оборотов на отрезке ДНК) и блокирует дальнейшее расплетание двойной спирали. Топоизомеразы решают названную топологическую проблему путем внесения одноцепочечных (топоизомераза I) или двухцепочечных (топоизомераза II) разрывов с последующим их восстановлением (рис. 25).

Рис. 25. Действие топоизомеразы I
2. Ферменты полимеризации
а) ДНК-праймаза синтезирует короткую РНК-затравку (праймер). ДНК-праймаза может быть отдельным ферментом (как у бактерий) или входить в качестве субъединицы в ДНК-полимеразу (как у α-ДНК-полимеразы эукариот). У некоторых прокариот (E. coli) праймаза образует комплекс с хеликазой, называемый праймосомой. Праймосома в свою очередь является компонентом еще более сложного комплекса – реплисомы, осуществляющей процесс полной репликации.
б) ДНК-полимеразы осуществляют комплементарное копирование одноцепочечной родительской ДНК. Про- и эукариотические ДНК-полимеразы несколько различаются по физическим и ферментативным свойствам, однако механизм их действия в общих чертах одинаков.
ДНК полимеразы прокариот разделяют на три типа – I, II и III (названия даны в порядке их открытия).
ДНК-полимераза I состоит из одного полипептида (молекулярная масса 109 кДа) и характеризуется:
- полимеразной активностью, т. е. осуществляет присоединение очередных нуклеотидов к строящейся цепи ДНК;
- 3′-экзонуклеазной (корректирующей) активностью, которая осуществляется в тех случаях, когда произошла ошибка и в строящуюся цепь включен «неправильный» нуклеотид. Тогда фермент отщепляет с растущего 3′-конца последний нуклеотид, после чего полимеризация восстанавливается;
- 5′-экзонуклеазной активностью, т. е. осуществляет последовательное отщепление нуклеотидов с 5′-конца РНК-затравки предшествующего фрагмента (рис. 26). Функция ДНК-полимеразы I исчерпывается, когда растущий фрагмент доходит до дезоксирибонуклеотидов предыдущего фрагмента.

Рис. 26. ДНК-полимераза на «стыке» синтезируемых фрагментов
Фактически ДНК-полимераза I – это два фермента на одной полипептидной цепи: при мягком расщеплении этой ДНК-полимеразы трипсином можно получить большой и малый фрагменты с разными активностями. Большой фрагмент ДНК-полимеразы I (фрагмент Кленова) обладает полимеразной и 3'-экзонуклеазной (корректирующей) активностями, малый фрагмент несет 5'-экзонуклеазную активность (рис. 27). Подобные свойства ДНК-полимеразы I соответствуют ее функциям в клетке: эта полимераза удаляет различного рода дефекты в ДНК в ходе репарации и служит вспомогательной полимеразой при репликации ДНК, удаляя РНК-затравки. В целом ДНК-полимераза I имеет большее отношение к созреванию реплицирующейся ДНК, чем непосредственно к полимеразным процессам в репликативной вилке.

Рис. 27. Фрагменты полимеразы I
ДНК-полимераза II – одиночный полипептид с молекулярной массой около 120 кДа, обладающий полимеразной и 3'-экзонуклеазной активностями. Его содержание в клетке в несколько раз ниже, чемДНК-полимеразы I. Эта полимераза лучше всего работает на двуцепочечной ДНК с одноцепочечными брешами в несколько десятков нуклеотидов длиной. Предполагают, что основной функцией ДНК-полимеразы II является достраивание поврежденных участков в молекуле ДНК, т. е. репарация ДНК.
ДНК-полимераза III играет главную роль в репликации ДНК у бактерий. В каждой клетке содержится только 10-20 копий фермента, примерно столько же, сколько репликативных вилок. ДНК-полимераза III состоит из нескольких субъединиц (α, β, γ, δ, δ′, ε, θ и т. д. – всего около 10) и обладает полимеразной и 3'-экзонуклеазной активностями. Репликацию проводит полная форма фермента – холофермент, содержащий все субъединицы. Полимеразную реакцию осуществляет каталитический кор (ядро) из α-, ε-, θ-субъединиц, остальные усиливают его действие. Отличительная черта ДНК-полимеразы III – исключительно высокая процессивность (скорость и точность синтеза новой цепи).
В клетках эукариотических организмов обнаружены шесть ДНК-полимераз: α, β, δ, ε, γ, ζ, из них первые четыре непосредственно участвуют в репликации хромосомной ДНК.
3. Ферменты, завершающие репликацию ДНК
а) ДНК-лигаза осуществляет «сшивание» соседних фрагментов ДНК, образуя межнуклеотидную (фосфодиэфирную связь). Реакция сопряжена с гидролизом молекулы АТФ.
б) Теломераза участвует в репликации теломерных (концевых) участков линейной ДНК.
ДНК-полимеразная система, описанная выше, оставляет недореплицированными 3′-концы материнских цепей ДНК, т. е. новые цепи оказываются укороченными с 5′-концов (рис. 28). Пропуск, образовавшийся после удаления крайнего праймера на 5′-конце дочерней цепи ДНК, заполнен быть не может, поскольку любая из полимераз не способна действовать «с нуля», а лишь удлиняет 3′-конец имеющегося полинуклеотида. В результате выступающие 3′-концевые участки материнской цепи ДНК остаются однотяжевыми, недореплицированными. Подобные концы ДНК называют острыми, или оверхенгами. Каждый раунд репликации ДНК будет приводить к ее укорочению на 50-60 нуклеотидов.
На концах линейных хромосом эукариот находятся специализированные ДНК-белковые структуры – теломеры. У большинства организмов теломерная ДНК представлена многочисленными короткими повторами (у млекопитающих – ТТАГГГ), не несущими генетической информации. Поэтому, если происходит потеря некоторой части данных повторов, это не отражается на функционировании генома. В этом состоит, вероятно, основная функция теломер: они предохраняют от недорепликации более значимые области ДНК, т. е. выполняют роль своеобразного буфера.
Однако теломерные участки ДНК имеют и ряд специальных функций (участвуют в фиксации хромосом к ядерному матриксу, влияют на экспрессию генов и др.), поэтому их укорочение допустимо лишь до некоторого предела. Синтез теломерных участков, утраченных в результате «концевой недорепликации», осуществляет фермент теломераза. К 3′-концу родительской цепи теломераза последовательно достраивает несколько десятков или сотен повторов ТТАГГГ. Таким образом, фермент удлиняет не дочернюю укороченную цепь, а родительскую – более длинную (рис. 28). Роль матрицы для наращивания ДНК повторами выполняет РНК, связанная с теломеразой. Следовательно, теломераза выступает как обратная транскриптаза – фермент, осуществляющий синтез ДНК на РНК-матрице.

Рис. 28. Схема репликации теломерного участка ДНК с участием теломеразы
Значительно удлиненная родительская цепь становится способной выступать в качестве матричной для образования еще одного фрагмента Оказаки дочерней цепи. При этом репликация идет в обычном порядке: праймаза синтезирует РНК-затравку, ДНК-полимераза последовательно присоединяет к затравке дезоксирибонуклеотиды – комплементарно теломерным повторам родительской цепи. Рост фрагмента происходит в обычном направлении 5′®3′. «Сшивание» фрагмента с цепью осуществляет ДНК-лигаза, экзонуклеаза удаляет РНК-затравку на новой цепи. В итоге конец двухцепочечной ДНК приобретает ту же конфигурацию, что и до действия теломеразы (дочерняя цепь немного короче старой), однако становится длинней на серию теломерных повторов. Это основной способ восстановления длины теломер, обнаруженный практически у всех исследованных организмов.
Высокая теломеразная активность наблюдается в эмбриональных тканях, стволовых и половых клетках (табл. 2). При дифференциации зародышевых или стволовых клеток активность теломеразы падает и теломеры укорачиваются. С достижением критической величины теломерной ДНК запускаются процессы остановки клеточного цикла. Дифференцированные клетки делятся ограниченное число раз. Это явление получило название «лимита Хейфлика».
Таблица 2
|
из
5.00
|
Обсуждение в статье: Ферменты, участвующие в репликации ДНК |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы