 |
Главная |


Схема электромеханической связи в мышечном волокне
|
из
5.00
|

А: состояние покоя, Б — возбуждение и сокращение
пд — потенциал действия, мм — мембрана мышечного волокна,
п – поперечные трубочки, т — продольные трубочки и цистерны с ионами Са 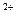 ,а — тонкие нити актина, м — толстые нити миозина с утолщениями (головками) на концах. Зет-мембранами ограничены саркомеры миофибрилл. Толстые стрелки — распространение потенциала действия при возбуждении волокна и перемещение ионов Са из цистерн и продольных трубочек в миофибриллы, где они содействуют образованию мостиков между нитями актином и миозином и скольжение этих нитей (сокращение волокна) за счет гребковых движений головок миозина.
,а — тонкие нити актина, м — толстые нити миозина с утолщениями (головками) на концах. Зет-мембранами ограничены саркомеры миофибрилл. Толстые стрелки — распространение потенциала действия при возбуждении волокна и перемещение ионов Са из цистерн и продольных трубочек в миофибриллы, где они содействуют образованию мостиков между нитями актином и миозином и скольжение этих нитей (сокращение волокна) за счет гребковых движений головок миозина.
Энергия гребкового движения одного мостика производит перемещение на 1 % длины актиновой нити. Для дальнейшего скольжения сократительных белков друг относительно друга мостики между актином и миозином должны распадаться и вновь образовываться на следующем центре связывания Са . Такой процесс происходит в результате активации в этот момент молекул миозина. Миозин приобретает свойства фермента АТФ-азы, который вызывает распад АТФ. Выделившаяся при распаде АТФ энергия приводит к разрушению имеющихся мостиков и образованию в присутствии Са новых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна — через 20 мс.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает поперечная исчерченность мышцы, видимая под микроскопом.
Расслабление мышечного волокна связано с работой особого механизма — «кальциевого насоса», который обеспечивает откачку ионов Са из миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
Сокращение мышц
Изучение работы мышцы с различными нагрузками и в разном темпе позволило вывести закон средних нагрузок и среднего темпа движений: максимальную механическую работу мышца совершает при средних нагрузках и среднем темпе движений. При высоких скоростях сокращения мышцы часть ее энергии тратится на преодоление сопротивления (растущего внутреннего трения и вязкости мышцы), а при низких скоростях — на поддержание изометрического напряжения, которое также присутствует в этом случае для закрепления достигнутой длины мышцы в каждый данный момент времени.
В зависимости от частоты нервных импульсов, поступающих от мотонейронов к мышечным волокнам, различают: одиночное и тетаническое сокращение мышц.
Одиночное сокращение мышечных волокон, возникающее в ответ на единичный импульс, поступающий из ЦНС по нервному волокну. В нем различают 3 фазы:
1. Латтентный (скрытый) период самый короткий. Начинается с момента раздражения и длится до начала сокращения (период электромеханической передачи).
2. Фаза сокращения.
3. Фаза расслабления обычно в 1,5-2 раза продолжительнее фазы сокращения, а при утомлении мышцы она значительно увеличивается.
Если раздражающие импульсы приходят к мышечному волокну один за другим и интервалы между ними короче, чем длительность одиночного сокращения, то возникает явление суперпозиции (суммации): механический эффект, когда отдельные сокращения мышечного волокна накладываются друг на друга, формируя при этом сложное сокращение - тетанус. Амплитуда тетануса больше величины одиночных сокращений.
Различают две формы тетануса:
1. Зубчатый тетанус возникает при более низкой частоте поступающих стимулов, когда каждый следующий нервный импульс поступает к мышечному волокну в фазу расслабления одиночных сокращений.
2. Гладкий (сплошной) тетанус возникает при более частом раздражении, когда каждый следующий импульс поступает в фазу сокращения мышечного волокна.
Сокращение целой мышцы зависит от количества одновременно работающих отдельных ДЕ и их координации во времени. Так при длительной, но не очень интенсивной работе отдельные ДЕ включаются в работу попеременно (бег на длинные и сверхдлинные дистанции). В этом случае ДЕ могут развивать как одиночные, так и тетанические сокращения в зависимости от частоты нервных импульсов.
При мощных кратковременных усилиях (поднятие штанги) активность отдельных ДЕ синхронизируется, т.е. одновременно возбуждаются почти все ДЕ работающей мышцы. При этом осуществляется мощное, но очень быстро приводящее к утомлению тетаническое сокращение.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа или с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных ДЕ. В случае же значительных напряжений потенциалы действия многих ДЕ алгебраически суммируются и возникает сложная интегрированная кривая записи электрической активности целой мышцы — электромиограмма (ЭМГ).
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе — вид отдельных пачек импульсов, которые совпадают с начальным моментом сокращения мышцы и разделены периодами перерывов. Особенно хорошо ритмичность появления подобных пачек наблюдается у спортсменов при циклической работе. У маленьких детей и неадаптированных к такой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
6. Структурные и физиологические основы мышечной силы
Движение является результатом взаимодействия внутренних и внешних сил, развиваемых в опорно-двигательном аппарате. Активные силы возникают при сокращении или напряжении мышцы во время ее возбуждения. Пассивные силы возникают при упругом напряжении, при растяжении мышцы, сопротивлении мышцы и ее сухожилия.
Сила мышцы зависит отряда морфологических и физиологических факторов: количества и свойств мышечных волокон в мышце, исходной длины мышцы, характера нервных импульсов, механических условий действия мышцы на кости скелета. Сила мышцы является суммой силы отдельных ее мышечных волокон.
Большое значение имеет анатомическое строение мышцы. В параллельно-волокнистых и веретенообразных мышцах (камбаловидная мышца и др.) сила мышц тем больше, чем больше ее анатомический поперечник, т. е. площадь поперечного сечения целой мышцы. В перистых мышцах (двуглавая мышца и др.) физиологический поперечник, т. е. площадь поперечного сечения всех мышечных волокон, гораздо больше, чем ее анатомический поперечник. В такой мышце содержится значительно больше мышечных волокон и, соответственно, больше ее сила.
На силу сокращения мышцы влияет ее исходная длина, так как от нее зависит возможное количество поперечных мостиков между актином и миозином. Предполагают, что в каждом цикле присоединения-отсоединения поперечных мостиков расходуется энергия 1 молекулы АТФ на 1 поперечный мостик. Следовательно, чем больше образуется в мышечном волокне актино-миозиновых мостиков, тем выше скорость расщепления АТФ, больше тяга сократительных белков и, соответственно, больше развиваемая мышцей сила.
Наибольшее количество актино-миозиновых контактов образуется при небольшом растяжении мышцы до некоторой оптимальной длины. При значительном растяжении саркомера нити актина далеко расходятся в стороны и практически не контактируют с расположенным в средней части саркомера миозином. В случае же резкого уменьшения длины саркомера нити актина в центре перекрывают друг друга, препятствуя контактам с миозином и также уменьшая число образуемых мостиков. В связи с этими особенностями взаимодействия сократительных белков наибольшая сила мышцы проявляется при некотором ее предварительном растяжении.
Одной из важнейших характеристик скелетных мышц, влияющих на силу сокращения, является состав мышечных волокон, т.е. их композиция.
Различают 3 типа мышечных волокон:
1. Медленные неутомляемые (I типа) — это выносливые (неутомляемые) и легковозбудимые волокна, с богатым кровоснабжением, большим количеством митохондрий, запасов миоглобина и с использованием окислительных процессов энергообразования (аэробные). Их обозначают также SO — Slow Oxidative (англ. — медленные окислительные). Их, в среднем, у человека 50%. Они легко включаются в работу при малейших напряжениях мышц, очень выносливы, но не обладают достаточной силой. Чаще всего они используются при поддержании ненагрузочной статической работы, например, при сохранении позы.
2. Быстрые неутомляемые или промежуточные (II-а типа) FG — Fast Glicolitic (быстрые гликолитические) используют анаэробные процессы энергообразования (гликолиз). Они менее возбудимы, включаются при больших нагрузках и обеспечивают быстрые и мощные сокращения мышц. Зато эти волокна быстро утомляются. Их примерно 30%. Волокна промежуточного типа (II-а) — быстрые неутомляемые, окислительные, их около 20%. В среднем, для разных мышц характерно различное соотношение медленных неутомляемых и быстрых утомляемых волокон. Так, в трехглавой мышце плеча преобладают быстрые волокна (67%) над медленными (33%), что обеспечивает скоростно-силовые возможности этой мышцы, а для более медленной и выносливой камбаловидной мышцы характерно наличие 84% медленных и всего 16% быстрых волокон.
3. Быстрые утомляемые (II-б типа).
Cостав мышечных волокон одной и той же мышцы имеет большие индивидуальные различия, которые зависят от врожденных типологических особенностей человека. К моменту рождения человека его мышцы содержат лишь медленные волокна, но под влиянием нервной регуляции устанавливается в ходе онтогенеза генетически заданное индивидуальное соотношение мышечных волокон разного типа. По мере перехода от зрелого возраста к пожилому число быстрых волокон у человека заметно снижается и, соответственно, уменьшается мышечная сила.
Количество тех или других мышечных волокон не изменяется в процессе тренировки. Возможно только нарастание толщины (гипертрофия) отдельных волокон, а также некоторое изменение свойств промежуточных волокон. При направленности тренировочного процесса на развитие силы происходит нарастание объема быстрых волокон, что и обеспечивает повышение силы тренируемых мышц.
Характер нервных импульсов изменяет силу сокращения мышц тремя способами:
1) увеличением числа активных ДЕ — это механизм вовлечения или рекрутирования ДЕ (сначала происходит вовлечение медленных и более возбудимых ДЕ, затем — высокопороговых быстрых Д Е);
2) увеличением частоты нервных импульсов, в результате чего происходит переход от слабых одиночных сокращений к сильным тетаническим сокращениям мышечных волокон;
3) увеличением синхронизации ДЕ, при этом происходит увеличение силы сокращения целой мышцы за счет одновременной тяги всех активных мышечных волокон.
Существенное значение имеют механические условия работы мышцы —точка приложения ее силы и точка приложения сопротивления (поднимаемого груза). Например, при сгибании в локте вес поднимаемого груза может быть порядка 40 кг и более, при этом сила мышц-сгибателей достигает 250 кг, а тяга сухожилий — 500 кг.
Между силой и скоростью сокращения мышцы существует определенное соотношение, имеющее вид гиперболы (соотношение сила — скорость, по А. Хиллу). Чем выше сила, развиваемая мышцей, тем меньше скорость ее сокращения, и, наоборот, с нарастанием скорости сокращения падает величина усилия. Наибольшую скорость развивает мышца, работающая без нагрузки. Скорость мышечного сокращения зависит от скорости передвижения поперечных мостиков, т. е. от частоты гребковых движений в единицу времени. В быстрых ДЕ эта частота выше, чем в медленных ДЕ, и, соответственно, потребляется больше энергии АТФ. Во время сокращения мышечных волокон в 1 с происходит примерно от 5 до 50 циклов прикрепления-отсоединения поперечных мостиков. При этом никаких колебаний силы в целой мышце не ощущается, так как ДЕ работают асинхронно. Лишь при утомлении возникает синхронная работа ДЕ, и в мышцах появляется дрожь (тремор утомления).
7. Работа мышцы
Механическая работа (А), совершаемая мышцей, измеряется произведением поднимаемого веса (Р) на расстояние (h): А = P*h кгс/м. При регистрации работы изолированной мышцы лягушки видно, что чем больше величина груза, тем меньше высота, на которую его поднимает мышца. Различают 3 режима работы мышцы:
1. Изотонический режим, или режим постоянного тонуса мышцы, наблюдается при отсутствии нагрузки на мышцу, когда мышца закреплена с одного конца и свободно сокращается. Напряжение в ней при этом не изменяется. Это происходит при раздражении изолированной мышцы лягушки, закрепленной одним концом на штативе. Так как при этих условиях Р = 0, то механическая работа мышцы также равна нулю (А = 0). В таком режиме работает в организме человека только одна мышца — мышца языка. В физиологической литературе часто встречается термин «изотонический режим» по отношению к такому сокращению мышцы с нагрузкой, когда по мере изменения длины мышцы ее напряжение сохраняется неизменным. Однако в этом случае механическая работа мышцы не равна нулю, т. е. она совершает внешнюю работу.
2. Изометрический режим - это режим постоянной длины мышцы, или статическая форма сокращения. Он характеризуется напряжением мышцы в условиях, когда она закреплена с обоих концов или когда мышца не может поднять слишком большой груз. При этом h = 0 и, соответственно, механическая работа тоже равна нулю (А = 0). Этот режим наблюдается при сохранении заданной позы и при выполнении статической работы. В этом случае в мышечном волокне все равно происходят процессы возникновения и разрушения мостиков между актином и миозином, т. е. тратится энергия на эти процессы, но отсутствует механическая реакция перемещения нитей актина вдоль миозина. Физиологическая характеристика такой работы заключается в оценке величины нагрузки и длительности работы: А=P*t, кг/мин.
3. Ауксотонический режим (смешанный режим) характеризуется изменением длины и напряжения мышцы, при сокращении которой происходит перемещение груза. В этом случае совершается механическая работа мышцы (А= P*h ). Такой режим проявляется при выполнении динамической работы мышц даже при отсутствии внешнего груза, так как мышцы преодолевают силу тяжести, действующую на тело человека. Различают 2 разновидности этого режима работы мышц: преодолевающий (концентрический) и уступающий (эксцентрический) режим.
|
из
5.00
|
Обсуждение в статье: Схема электромеханической связи в мышечном волокне |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы