 |
Главная |


Отдел Оомикота – Oomycota
|
из
5.00
|
Этот отдел объединяет организмы у которых зооспоры двужгутиковые с гетероморфными и гетероконтными жгутиками (передний перистый, задний гладкий). Бесполое размножение при помощи двужгутиковых зооспор, у наземных форм – конидий. Половой процесс оогамия, завершающийся формированием ооспоры. Вегетативное тело – хорошо развитый неклеточный диплоидный мицелий. В оболочках клеток содержится целлюлоза и редко небольшое количество хитина (Leptomitales). В качестве запасного продукта накапливается крахмал и водорастворимый глюкан – миколаминарин. Синтез лизина осуществляется по типу, сходному для всех автотрофных растений – через диаминопимелиновую кислоту. У оомицетов обнаружен очень редкий фермент – глюказа, легко разлагающий молочную кислоту. Митохондрии имеют трубчатую структуру крист.
В отделе один класс Oomycetes, в котором выделяют 4 порядка различающиеся, прежде всего бесполым размножением.
Порядок Lagenidiales. Водные паразиты. Вегетативное тело в виде мешковидной, слабо разветвленной гифы, формирующей зооспорангии, оогонии и антеридии.
Порядок Leptomitales. Водные сапротрофы. Вегетативное тело в виде тонкого нитчатого разветвленного мицелия с перетяжками — ложными перегородками. Зооспорангии развиваются либо на верхушке гифы, либо интеркалярно.
Порядок Saprolegniales. Водные сапротрофы и паразиты. Мицелий хорошо развит, разветвленный ценотический. Спорангии, хотя и отделены от несущих гиф септами, выглядят просто вздутыми участками. Монопланетические и дипланетические формы. В оогонии несколько яйцеклеток и соответственно формируется несколько ооспор.
Порядок Peronosporales. Мицелий либо поверхностный, либо межклеточный, последний формирует гаустории, внедряющиеся в клетки-хозяина. В пределах порядка прослеживается переход от сапротрофов к паразитам и от водного образа жизни к наземному существованию. Бесполое размножение осуществляется двужгутиковыми зооспорами или у высокоорганизованных представителей – конидиями. Половой процесс – оогамия, которая протекает чаще всего в межклетниках растения-хозяина. В оогонии, как правило, одна яйцеклетка и соответственно одна ооспора.
Отдел Oomycota в своем происхождении связывают с разножгутиковыми желто-зелеными водорослями, о чем свидетельствует ряд морфологических (сходство в строении таллома и жгутикового аппарата), биохимических (сходство пути синтеза лизина, состав полисахаридов клеточной стенки) и цитологических особенностей (строение крист митохондрий – у оомицетов они имеют трубчатую структуру, а не пластинчатую, как у остальных групп грибов).
Порядок сапролегниевые – Saprolegniales
Преимущественно сапротрофы, развиваются на органике животного происхождения: на трупах беспозвоночных, икре рыб и лягушек. Иногда они поселяются на ослабленных рыбах и мальках, вызывая их массовую гибель.
У сапролегниевых хорошо развит неклеточный вегетативный мицелий, часто многократно разветвленный, многоядерный. Поперечные перегородки возникают при образовании органов бесполого и полового размножения. Цилиндрические зооспорангии развиваются на концах вегетативных гиф. У одних видов зооспорангии одиночные, у других – образуется группа зооспорангиев в виде кустиков. У форм с одиночными зооспорангиями можно наблюдать врастание нового зооспорангия в оболочку старого, опустевшего. В опустевший зооспорангии поступает новая часть протоплазмы, которая одевается внутри него собственной оболочкой. Это явление получило название пролиферации зооспорангиев. У сапролегниевых грибов наблюдается два типа зооспор – грушевидные с двумя жгутиками на переднем конце зооспоры и бобовидные с боковым расположением жгутиков. Грушевидные зооспоры некоторое время плавают, затем останавливаются, одеваются оболочкой и из внутренней части успокоившихся спор выходят новые зооспоры уже бобовидной формы с двумя расположенными сбоку (у вогнутой части) жгутиками. Это явление называется дипланетизм, а зооспоры называются диморфные.
В пределах порядка Saprolegniales у разных родов встречаются зооспоры различного типа (рис. 5). У Saprolegnia из зооспорангия выходят грушевидные зооспоры. Прорастание таких вторичных зооспор осуществляется после того, как они приобретут оболочку. У видов Pythiopsis встречаются только зооспоры грушевидной формы, зооспоры бобовидные здесь отсутствуют. У представителей p. Achlya имеются только бобовидные зооспоры. Виды Pythiopsis и Achlya относятся к монопланетическим формам, а зооспоры их мономорфные. У представителей Aplones подвижные зооспоры вообще не выходят из зооспорангия. Прорастание их происходит внутри общей оболочки зооспорангия. Зрелый зооспорангии выглядит щетинистым.
Таким образом, у сапролегниевых можно проследить смену дипланетических зооспор (диморфных) неподвижными спорами, что является прогрессивным направлением развития, дающим возможность приспосабливаться к наземному существованию.
Взаимное расположение оогониев и антеридиев у представителей данного порядка может быть различным. Наиболее часто встречается случай, когда на одной и той же ветви грибницы формируются оогоний и антеридий. У других видов оогоний и антеридии возникают далеко друг от друга: антеридий на одних ветвях, оогоний — на других. Известны также случаи полного разделения полов – на одном мицелии развиваются оогоний, на другом — антеридии (гетероталличные формы).
Перед оплодотворением в оогонии происходит уменьшение числа ядер. Вокруг оставшихся ядер обособляется протоплазма и формируется несколько яйцеклеток (оосфер). В антеридии к этому времени образуются выросты – полинодии, через которые содержимое антеридия передается в оогоний к яйцеклеткам, через поры в оболочке оогония. Образовавшиеся зиготы покрываются оболочкой и превращаются в ооспоры, которые зимуют. Весной ооспоры прорастают в короткую гифу с зооспорангием на конце.
У сапролегниевых нередко наблюдается партеногенез — образование ооспор без оплодотворения, при полном отсутствии антеридия. В этих случаях ооспоры являются гаплоидными.
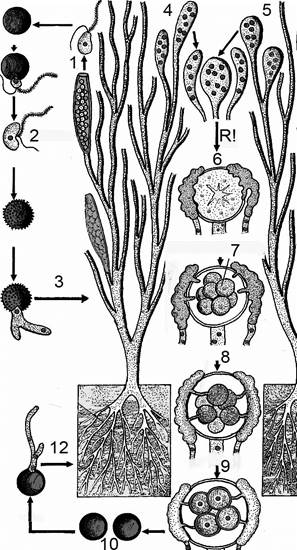
| Рис. 5. Цикл развития Saprolegnia. 1 – выход грушевидной зооспоры из зооспорангия 2 – бобовидная зооспора 3 – прорастание гифы в мицелий 4 – антеридий 5 – оогоний 6 – образование гаплоидных ядер 7 – оосфера 8 – гаметангиогамия 9 – формирование ооспор 10 – ооспора 12 – прорастание ооспоры в мицелий |
Порядок пероноспоровые – Реrоnosроrales
Грибы с хорошо развитым неклеточным мицелием, которые в большинстве случаев ведут паразитический образ жизни на высших растениях. Среди них есть и сапротрофы, поселяющиеся на мертвых растительных и животных остатках, находящихся в воде.
У наиболее примитивных представителей (Pythiaceae) дифференциации мицелия, как правило, не наблюдается. Настоящие гаустории на гифах мицелия имеются, главным образом, у высокоорганизованных представителей (Peronosporaceae, А1buginaceae), тогда как у Phytophthoraceae они еще слабо выражены или отсутствуют.
Бесполое размножение осуществляется с помощью зооспор или конидий. Зооспорангии – овальные, яйцевидные или шаровидные. Зооспоры – почковидные с двумя боковыми жгутиками, перистый направлен вперед, гладкий – назад. Конидии, у большинства видов, образуются на ветвях спорангиеносца (конидиеносца) одиночно, иногда цепочками (Albuginaceae). Спорангиеносцы могут быть разных типов: малодифференцированными, напоминающими гифы мицелия и способными после образования конидий (зооспорангиев) развиваться дальше и давать новые разветвления (Pythiaceae и Phytophthoraceae); но чаще всего они хорошо обособлены от мицелия, имеют свой тип ветвления и не способны к дальнейшему росту. Такие спорангиеносцы (конидиеносцы) выступают наружу через устьица. Спорангиеносцы у видов Albuginaceae развиваются под эпидермисом растения в виде ложа, как правило, они неразветвленные, короткие, булавовидной формы.
Прорастание спор протекает по-разному: либо дифференцируя свое содержимое, они дают начало зооспорам, которые активно передвигаются в воде, и затем каждая формирует мицелиальный росток, либо сразу образуют мицелиальный росток, которым и осуществляется новое заражение. Первый тип прорастания наблюдается у филогенетически более древних представителей порядка (Pythiaceae, частично Phytophthoraceae, а также Albuginaceae), тогда как второй тип характерен для филогенетически более молодых его представителей (Peronospora, частично у Phytophthora). У Phytophthora infestans, в зависимости от условий окружающей среды прорастание может осуществлятся двумя способами.
Встречается и третий тип прорастания, когда содержимое споры выходит наружу в виде голого протоплазматического пузыря, и затем в нем формируются зооспоры (Pythiaceae, Peronosporaceae).
Половой процесс – оогамия. Обычно внутри тканей растения формируется крупный шаровидный оогоний и на том же мицелии возникают удлиненные антеридии. В каждом оогонии формируется одна или несколько оосфера, которые после оплодотворения одеваются многими оболочками и превращаются в ооспоры, наружные оболочки которых могут иметь гладкую, бугорчатую или сетчатую поверхность.
Прорастает ооспора чаще всего после перезимовки, либо с образованием тонкостенного зооспорангия, либо спорангиеносца (конидиеносца) с зооспорангиями (конидиями), либо сразу мицелиальным ростком, внедряющимся в ткань питающего растения.
Особенностью большинства переноспоровых является их узкая специализация, при которой каждый вид гриба может развиваться только на одном виде растения-хозяин (Sclerospora поражает злаки, Plasmopara – зонтичные и виноградовые, Albugo – крестоцветные), хотя встречаются виды и с широкой специализацией (Pythium паразитирует на корнях сеянцев древесных пород и рассаде свеклы, капусты, огурца, табака, кукурузы). Представители пероноспоровых вызывают гниение пораженных тканей, образование пятен на пораженных листьях, сопровождающихся появлением налета белого, серого или фиолетового цвета (ложная мучнистая роса), а для некоторых представителей характерно (Albuginaceae) образование белых выпуклых пустул (белая ржавчина), что влечет за собой отмирание и гибель пораженной ткани и органа растения.
В порядке четко выделяются две эволюционные тенденции:
1. Выход организмов из водной среды к наземному существованию, при этом зооспоры сменяются конидиями. Наземные виды выработали гигроскопические торзионные механизмы распространения спор. Древовидные спорангиеносцы под влиянием небольшого изменения влажности воздуха способны к вращению вокруг длинной оси, при этом созревшие конидии сбрасываются с ветвей и подхватываются токами воздуха.
2. Совершенствование облигатного паразитизма с узкой специализацией отдельных видов. Возникают гаустории, открытое спороношение сменяется закрытым – спорангиеносцы развиваются не на поверхности листа, а между эпидермисом и кутикулой, используя последнюю как барьер, защищающий гриб от внешних воздействий.
4.Особенности биологии Phytophthora infestans
Немецкий ботаник Антон де Бари, описавший возбудителя фитофтороза картофеля, дал ему имя Phytophthora infestans,что означает – инфекционный пожиратель растений.
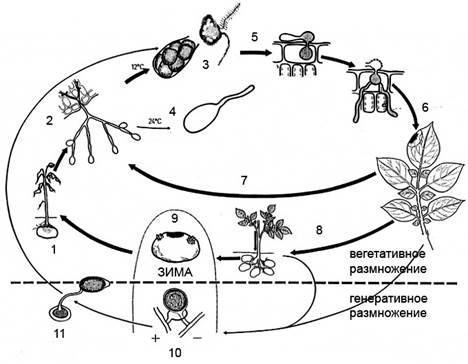
Мицелий фитофторы – бесцветный неклеточный разветвленный, погруженный в ткань растения-хозяина. Мицелий распространяется по межклетникам, проникает внутрь клеток гаусториями. На нижней стороне листа, через разрывы пораженной ткани или через устьица, высовываются разветвленные спорангиеносцы с лимоновидные зооспорангиями (рис. 6), которые разносятся дождевыми брызгами или ветром. Попадая в каплю воды, зооспорангий раскалывается, образуется шесть-восемь двужгутиковых зооспор, покидающих зооспорангий через отверстие на вершине. Проплавав некоторое время, зооспора теряет жгутики, покрывается оболочкой (инцистируется) и прорастает ростковой трубкой, которая внедряется в ткань листа или плода. Сигналом для инцистирования служит контакт с твердой поверхностью. При повышенной температуре (выше 20°) зооспоры не формируются, и зооспорангий прорастает целиком, как одна спора – конидия. В теплую погоду капли на листьях быстро высыхают и лишенные клеточной стенки зооспоры могут погибнуть. Так, ценой снижения потомства, повышается устойчивость развития и зооспорангий превращается в конидию.
|
| Рис. 6. Цикл развития Phytophthora infestans. 1 – развитие растения из пораженного клубня; 2 – спороношение; 3 – формирование зооспор; 4 – прорастание конидии; 5-6 – прорастание зооспоры или конидии и поражение растения; 8 – заражение клубня; 9 – пораженный клубень; 10 – половой процесс; 11 – прорастание зимующей ооспоры и заражение клубня. |
При благоприятных условиях (высокой влажности и умеренных температурах) через три-четыре дня после заражения на листе образуется новое спороношение. Единичное заражение дает несколько десятков тысяч спорангиев, прорастающих еще шестью-восемью зооспорами. Это обусловливает взрывной характер развития болезни, при благоприятной погоде за несколько недель поражаются все растения на поле. Споры с дождевыми токами проникают в почву и заражают формирующиеся клубни. Особенно сильно это происходит при уборке картофеля, когда поврежденные клубни контактируют с зараженной ботвой. Поражение клубней опасно не столько само по себе (при нормальном хранении фитофтора не переходит на здоровые), сколько снижением общего иммунитета. Фитофтора выделяет в зараженные клубни специфический глюкан, который подавляет образование раневой перидермы и защитных веществ. В результате такие клубни становятся легкой добычей грибов и бактерий, вызывающих быстрое гниение клубня и распространяющихся на соседние здоровые клубни. Партии картофеля, где более 10% зараженных фитофторой клубней, могут сгнить полностью.
Большинство видов фитофторы имеют в жизненном цикле половой процесс – оогамию. У гомоталличных видов мужские и женские структуры (антеридии и оогонии) формируются на одном мицелии, т.е. они обоеполы. У гетероталлических видов для полового процесса необходимо слияние оогониев и антеридиев, образованных генетически различными мицелиями (А1 и А2). В результате полового процесса появляется зигота - ооспора, покрытая толстой оболочкой и способная переносить неблагоприятные внешние условия (например, зимовку). У картофельной фитофторы ооспоры находили крайне редко. Весь ее цикл проходит в вегетативной фазе, а зимовка осуществляется в комфортных условиях хранилища. Единственным источником весеннего возобновления фитофторы считали зараженные клубни, т.к. европейские штаммы фитофторы только мицелий А1 и поэтому стерильны. Однако сравнительно недавно, в 80-х годах, в ряде европейских стран, в том числе и в России, обнаружили штаммы с типом спаривания А2 и многочисленные ооспоры. Ооспоры могут возникать в ткани пораженных томатов, зимовать в растительных остатках и заражать весной и томаты, и картофель. Вполне возможно это связано с вывозом семенного картофеля из Мексики, который обеспечил распространение мексиканских штаммов фитофторы по всему миру. Это привело, к появлению типа спаривания А2, возможности полового процесса и зимовки ооспор в почве (вне клубней).
|
из
5.00
|
Обсуждение в статье: Отдел Оомикота – Oomycota |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы