 |
Главная |


КОНЦЕНТРАЦИИ ВОЗБУЖДЕНИЯ
|
из
5.00
|
Многие нейроны оказывают свое воздействие на один и тот же нейрон (имеет место схождение потоков импульсов к одному и тому же нейрону). Шеррингтон называл это "принцип общего конечного пути". Например, сокращение мышцы (за счет возбуждения α‑мотонейрона) можно вызвать путем растяжения этой мышцы (рефлекс мышечных веретен), путем раздражения кожных рецепторов (сгибательный рефлекс) и т.п. Ч.Шеррингтон определяет физиологический механизм принципа «общего конечного пути» на уровне эфферентного звена рефлекторной дуги.
В основе концентрации лежит конвергенция (лат. Convergentio) возбуждения. Convergentio от Convergo Convergere – сближать, сходиться.
Конвергенция – способность двух или нескольких нейронов устанавливать синаптические связи с одним и тем же нейроном. [27]
Нервные центры высших отделов мозга являются мощными коллекторами, собирающими разнородную афферентную информацию. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. На это указывают прямые исследования центральных нейронов: в нервном центре имеется значительное количество поливалентных, полисенсорных нервных клеток, реагирующих на разномодальные и разновалентные стимулы (свет, звук, механические раздражения и т.д.). Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т.е. высокий уровень интеграционных функций.
Различают конвергенцию нервных импульсов мультисенсорную (говорилось выше), мультибиологическую, сенсорно-биологическую и эфферентно-афферентную.
Мультибиологическая конвергенция – схождение к одному нейрону двух или нескольких возбуждений от биологических раздражителей (холод, боль, жажда, половое влечение и т.п.)
Сенсорно-биологическая конвергенция – схождение к одному нейрону двух или нескольких возбуждений от сенсорных и биологических раздражителей (холод, боль, жажда, половое влечение и т.п.). Это один из механизмов обучения, условных рефлексов и афферентного синтеза функциональных систем.
Эфферентно-афферентная конвергенция или афферентно-эфферентная (синоним) наблюдается когда эфферентное возбуждение отходит от нейрона и взаимодействует с афферентным возбуждением, приходящим к нейрону в этот момент. Этот тип конвергенции является одним из механизмов акцептора результата действия.
Окклюзия
Окклюзия — взаимодействие двух импульсных потоков между собой. Впервые явление окклюзии было описано Ч.Шеррингтоном. Сущность его заключается во взаимном угнетении рефлекторных реакций, при котором суммарный результат оказывается значительно меньше, чем сумма взаимодействующих реакций
Окклюзия (occlusio; лат. occludo, occlusum закрывать, замыкать)
Синоним – заклинивание.


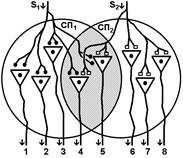
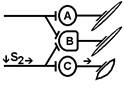
При последовательном раздражении
S1 + S2 = 2 +2 = 4

При одновременном раздражении
S’1 + S’2 = 3
S1 + S2 > S’1 + S’2
Согласно Ч.Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями взаимодействующих рефлексов. В связи с этим при одновременном поступлении двух афферентных посылок возбуждающий постсинаптический потенциал вызывается каждым из них отчасти в одних и тех же мотонейронах спинного мозга. Окклюзию используют в электрофизиологических экспериментах для определения общего звена для двух путей распространения импульсов. Если имеется общее звено, то одновременное раздражение двух путей с максимальной интенсивностью вызывает ответ меньшей величины, чем сумма ответов, получаемых при раздельном раздражении этих путей стимулами той же интенсивности.
Приведем два неточных объяснения явления окклюзии.
Первое. «Если два нервных центра рефлекторных реакций имеют частично перекрываемые рецептивные поля, то при совместном раздражении обоих рецептивных полей реакция будет меньше, чем арифметическая сумма реакций при изолированном раздражении каждого из рецептивных полей — феномен окклюзии» [28]
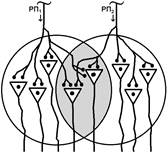
При объяснении явления окклюзии нужно говорить о перекрытии синаптических полей нервных центров, а не рецептивных полей.

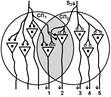
Второе. «За счет явления дивергенции один и тот же нейрон может передавать сигналы на ряд других нейронов, в результате чего возникает определенный эффект (например, происходит активация 10 мышечных волокон в мышце, в результате чего мышца развивает напряжение, равное 100 мгс. Второй нейрон возбуждает тоже 10 других волокон (100 мгс). Но если оба нейрона возбуждать одновременно, то суммарная активность мышцы будет 180 мг. Почему? Оказывается, часть волокон у них были общими (т. е. нейрон 1 и 2 передавали информацию на одни и те же волокна). Вот это явление получило название окклюзии или закупорки.»
Почему это трудно признать правильным объяснением? Нейроны не конвергируют на мышце.
В основе окклюзии лежит явление конвергенции.
Облегчение
В ряде случаев вместо такого ослабления реакции при совместном раздражении рецептивных полей двух рефлексов можно наблюдать феномен облегчения (т.е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Это результат того, что часть общих для обоих рефлексов нейронов при изолированном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций.
Синоним — суммация. Да да суммация. Какая? Пространственная.
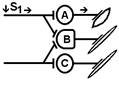
При последовательном раздражении
S1 + S2 = 1 +1 = 2
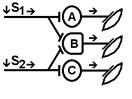
При одновременном раздражении
S’1 + S’2 = 3
S1 + S2 < S’1 + S’2
Последействие
Известно, что длительность ответной реакции не соответствует времени действия раздражителя и может развиваться и осуществляться через достаточно большой промежуток времени после завершения раздражения.
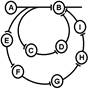
Один из вариантов этого свойства - длительное циркулирование импульсов по "нейронной ловушке". Итальянский физиолог Лоренто де Но обнаружил это явление: поступивший импульс может минутами или часами пробегать небольшой отрезок нейронной цепи. Благодаря этому, как полагают некоторые авторы, происходит перевод следа (энграммы) из краткосрочной памяти в долгосрочную.
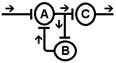
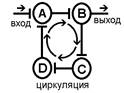
Рис. Реверберация возбуждения в нервной сети (по Лоренто де Но)
Предполагается, что развитие послеразрядной активности нейрона связано с особенностями проведения возбуждения через синапсы. Так как передача возбуждения в синапсах осуществляется с помощью медиаторов (в мотонейронах процесс эмиссии медиатора длится до 10 и более мс), сдвиг постсинаптического потенциала также имеет большую продолжительность. В случае развития ВПСП нейрон может разряжаться на протяжении длительного времени. Такая ритмическая импульсация затухает постепенно или обрывается внезапно.
Возможен третий механизм длительного последействия, который получил название пролонгирования возбуждения. В нервных центрах существуют цепочки нейронов, связанные между собой таким образом, что импульсация определенного нейрона может вызывать генерацию импульсов в других нейронах и от них вновь приходит к первому нейрону. При этом создается возможность длительной циркуляции нервной импульсации по длинным кольцевым связям и пролонгировать возбуждение в нервном центре.
Пролонгирование импульсации имеет очень большое значение в процессах переработки информации и особенно в фиксировании следов информации, т. е. памяти.

ТРАНСФОРМАЦИЯ РИТМА ВОЗБУЖДЕНИЯ
(лат. transformatio преобразование, превращение)
Синоним понятия — трансформация частоты следования импульсов.
Определение понятия:
Трансформация ритма возбуждений — одно из свойств проведения возбуждения в нервной системе (нервном центре), заключающееся в способности нейрона изменять ритм приходящих импульсов.[29]
Существует мнение, что способность трансформировать ритм возбуждения есть только у нейрона, причём у сомы нейрона. А, например, у скелетного миоцита эта способность отсутствует. При этом следует помнить, что скелетный миоцит, как и другие эффекторные клетки, могут играть роль частотного фильтра, т.е воспринимать частоты возбуждения до какой-то предельной.
Например, поступает импульс, идущий с частотой 25 Гц, а нейрон в ответ на это, возбуждаясь, генерирует 50 Гц, или наоборот, поступает 100 Гц, а выходят 40 Гц.
Особенно четко проявляется трансформация ритма возбуждения при раздражении афферентного волокна одиночными импульсами. На такой импульс нейрон отвечает пачкой импульсов. [30]
Выделяют несколько вероятных механизмов трансформации ритма возбуждения. [31]
В ряде случаев трансформация ритма возбуждения обусловлена возникновением длительного возбуждающего постсинаптического потенциала, на фоне которого развивается несколько спайков. [32]

Этот механизм трансформации ритма возбуждения возникновением длительного возбуждающего постсинаптического потенциала, объясняет принцип кодирования «сила стимула кодируется частотой импульсов, а не амплитудой»

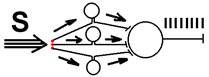
Другим механизмом[33] возникновения множественного разряда импульсов являются следовые колебания мембранного потенциала. Когда его величина достаточно велика, следовые колебания могут привести к достижению критического уровня деполяризации мембраны и обусловливают появление вторичных спайков. [34]
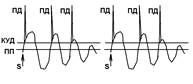
Как уже отмечалось выше способность трансформировать ритм возбудения приписывают только соме и отказывают в наличии этого свойства аксону. Аксону да, но не аксонному холмику.
От состояния аксонного холмика – порога раздражения – в значительной мере может зависеть направление (урежение или учащение) и выраженность трансформации частоты следования импульсов.

Временная суммация

!!! Говоря о трансформации ритма возбуждения лучше всё же говорить не об отдельной клетке (нейроне) или отдельной её части, а о модуле (колонке для коры), ансамбле нейронов, нерном центре.
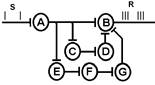

Каждый модуль, или нейронный ансамбль, представляет собой совокупность локальных нейронных сетей, которая обрабатывает информацию, передает ее со своего входа на выход, подвергает трансформации, определяемой общими свойствами структуры и ее внешними связями.[35]
Кодирование[36]
!!! Говоря о трансформации ритма возбуждения лучше всё же говорить не об изменении частоты, а об изменении паттерна.

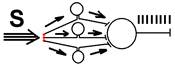

Принцип доминанты
Был открыт А.А.Ухтомским на основании опытов проведенных в 1904-1911 году.
Изучая ответы скелетной мышцы кошки на электрические раздражения коры больших полушарий, он обнаружил, что при акте дефекации ответы мышцы прекращаются. Проанализировав этот факт, Ухтомский пришел к мнению о наличии в ЦНС явления доминанты.
Речь идет о том, что среди рефлекторных актов, которые могут быть выполнены в данный момент времени, имеются рефлексы, выполнение которых представляет наибольший "интерес" для организма, они в данный момент времени самые важные. Поэтому эти рефлексы реализуются, а другие - менее важные - тормозятся. А.А.Ухтомский назвал центры, участвующие в реализации доминантных рефлексов - "доминантным очагом возбуждения".
"Доминантный очаг" обладает рядом важных свойств:
1. он стойкий (его сложно затормозить),
2. интенсивность его возбуждения усиливается слабыми раздражителями:
3. этот очаг тормозит другие потенциальные доминантные очаги.

Банунг (нем. Bahnung) — «проторение пути» суммационный рефлекс[37]
Инерционность доминанты обусловлена длительными следовыми процессами, механизмы которых детально освещены.[38] В естественных условиях длительное следовое возбуждение может быть обусловлено:
1. суммацией ВПСП приходящих подпороговых импульсов,
2. синаптической потенциацией (облегчением) при ритмическом раздражении пресинаптических входов
3. изменение концентрации К+ в снаптической щели, который как деполяризатор усиливает вхождение Ca++ в пресинаптическое окончание,
4. метаболическими следами, связанными с влиянием медиаторов на циклазные системы постсинаптических клеток,
5. циклическими связями в ЦНС, способными обеспечить следовую самостимуляцию центров.
Отчего же именно данный очаг возбуждения является доминантным? Это определяется состоянием организма, например, гормональным фоном. У голодного животного доминантными рефлексами являются пищевые. Развитием представления о доминанте являются работы Анохина о функциональной системе, в которой есть блок, принимающий решение. Именно на основе мотиваций и памяти (следов) происходит принятие решения ("Что делать в данный момент времени") с учетом, конечно, результатов афферентного синтеза. В настоящее время идет интенсивное изучение конкретных процессов, лежащих в основе становления и формирования доминантных очагов в ЦНС.
Доминанта как один из основных принципов координационной деятельности ЦНС имеет важное значение в жизни человека. Например, именно благодаря доминанте возможно сосредоточение психической (внимание) и выполнение умственной или физической деятельности (в данном случае - это трудовая доминанта). В период поиска пищи, поедания возникает пищевая доминанта. Существуют половая, оборонительная доминанта.
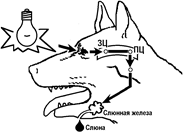
СИНАПТИЧЕСКАЯ ЗАДЕРЖКА
Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. [39] При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка).[40]
Синаптическая задержка — замедление скорости распространения возбуждения в синапсе (межклеточном контакте) относительно скорости распространеия возбуждения в клетках возбудимых тканей (нервных, мышечных). Относительно малую скорость распространения возбуждения в синапсе связывают с длительностью процессов выделения медиатора из пресинаптического окончания, диффузии его по синаптической щели и процесса взаимодействия его с постсинаптической мембраной. [41]
В нервных клетках высших животных и человека одна синаптическая задержка примерно равна 1 мс. [42]
Скорость этих процессов в 10 и более раз меньше, чем скорость распространения возбуждения по нерву. С. з. в химических синапсах обычно равна 0,2—0,5 мс.
Если учесть, что в реальных рефлекторных дугах имеются десятки последовательных синаптических контактов, становится понятной длительность большинства рефлекторных реакций — десятки миллисекунд. [43]
|
из
5.00
|
Обсуждение в статье: КОНЦЕНТРАЦИИ ВОЗБУЖДЕНИЯ |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы