 |
Главная |


Отбор быстрорастущих деревьев ели в возрасте 5, 7, 12, 30, 40 лет и формирование из них крупных деревьев (ретроспекция развития)
|
из
5.00
|
Вопрос диагностики роста дерева можно выяснить ретроспективным анализом ствола уже взрослого дерева вначале в самых простых сообществах древесных растений, какими являются лесные культуры. В качестве объекта для изучения желательны чистые высокопродуктивные лесные культуры, удовлетворяющие следующим требованиям:
- созданные одновозрастным посадочным материалом;
- с равномерным размещением посадочных мест;
- с максимальной сохранностью всех высаженных растений;
- в возрасте, приближающемся к возрасту рубки.
Такие культуры были обнаружены нами в 1983 г. в кв. 65 и 66 Сепычевского лесничества Сивинского лесхоза. По лесоустройству участок площадью 42 га считался посадками ели 1920 года. Однако изучение срубленных моделей показало, что на нем представлено два участка: основной массив культур 1913 года площадью 38 га и примыкающий к нему участок культур 1916 года площадью 4 га. Расположены они среди холмисто-увалистого ландшафта с лесистостью 50-60%, где вершины и склоны холмов заняты полями, а понижения и лога елово-пихтовым лесом. В 2,0 км на север от участка до 1986 года существовала д. Фарафоны, а в 1,3 км на юго-восток – д. Петрухино. В последней, по свидетельству ее жителя Носкова Филарета Викуловича, в 1930-40-х гг. насчитывалось 40 дворов. В настоящее время деревни брошены и ближайший населенный пункт – д. Катаевка (5,5 км) и д. Путино (10 км на восток).
На основном участке разместилось 600 рядов. Посадка проводилась 3-летними сеянцами по схеме 2,13×1,07 м (1 сажень на 0,5 сажени) и была настолько правильной, что спустя 70 лет прослеживались ряды не только в продольном и поперечном направлении, но и по двум диагоналям. Участок рубкам ухода не подвергался из-за труднодоступности –со стороны деревень дорога к нему проходила через заболоченные долины ручьев.
К основному массиву культур примыкали посадки, проведенные в 1916 г. на площади 4 га. Участок был дренированным, но плоским; при той же схеме посадки густота стояния живых растений оказалась меньшей (770 шт./га против 1200 шт./га). Вероятнее всего, после создания культур дополнения в них уже не проводили в связи с Первой мировой войной в отличие от основного участка, где такая возможность была.
Еще один участок культур ели площадью 10 га примыкал к основному с юга и был создан, по всем признакам, сплошным посевом семян по пашне в 1918 или в 1919 г. По-видимому, эта пашня была заброшена и оставшиеся в Очерском окружном лесничестве семена ели (возможно, многолетний страховой запас семян) были щедро здесь высеяны. Отсутствие прогалин и равномерная густота стояния деревьев никак не могли быть результатом естественного возобновления ели по пашне на такой большой территории, имеющей поперечник в 300 м; кроме того, в окружающих участок ельниках присутствовала пихта и густота в них была намного ниже и не была столь равномерной.
В этих культурах были заложены две пробные площади (табл. 5).
Таблица 5 – Характеристика пробных площадей в культурах ели в кв. 65 Сепычевского лесничества Сивинского лесхоза
| Показатели | Пробные площади | |
| Год создания | ||
| Возраст на момент изучения, лет | ||
| Способ создания | посадка | посев семян вразброс |
| Схема размещения посадочных мест, м | 2,13×1,07 | |
| Число растений, шт./га: | ||
| - в момент создания | ~12000 | |
| - в год учета, живых и мертвых | ||
| - в год учета только живых | ||
| Средний диаметр, см | 21,1 | 18,2 |
| Средняя высота, м | 22,2 | 20,7 |
| Класс бонитета | 1,6 | 1,6 |
| Средняя высота до живой кроны, м | 15,7 | 13,8 |
| Полнота абсолютная, м2/га | 43,6 | 50,7 |
| Полнота относительная по стандартным таблицам: - для естественных древостоев (по Разину, 1977, с. 437) - для лесных культур ( по Разин, Рогозин, 2011, см. табл. 3.4) | 1,03 0,70 | 1,25 0,84 |
| Запас древесины, м3/га |
После перечета деревьев было намечено 5 классов (рангов) диаметра, начиная с самых тонких живых (1-й ранг) и кончая самыми толстыми (5-й ранг). В каждый из них отбирали строго по 6 моделей. Поиск моделей имел важные особенности. Так, модели вблизи ПП 1 отбирали в возможно более плотном окружении, для чего вокруг модели закладывали пробную площадь с радиусом 3,2 м и учитывали диаметры живых, сухих деревьев и валежа. Условие отбора модели состояло в отсутствии пустых посадочных мест: в ряду более одного и в соседних рядах – более чем трех (из пяти). На втором участке пробы вокруг моделей не закладывали, но отбирали их с таким расчетом, чтобы на расстоянии 3-4 м от модели не встречались даже небольшие прогалины. При соблюдении этих условий на обоих участках отбирались деревья, которые с самого раннего возраста развивались в условиях конкуренции.
Перед рубкой на стволе дерева делали зарубки на высоте 30 и 130 см от почвы. Модель спиливали вначале на высоте 30 см, после чего разгребали подстилку, освобождая поверхность почвы, подрубали корневые лапы и делали нишу для введения шины бензопилы на уровне почвы. Затем спиливали пень на высоте 5 см от поверхности почвы. У модели измеряли общую высоту, высоту 10, 20 и 30 лет назад, высоту до живой кроны. После этого приступали к анализу поперечных спилов на высотах 5, 30 и 130 см. У средних и толстых моделей обычно удавалось снять все измерения на спилах прямо на месте с точностью до ±0,1 мм. У тонких деревьев измерения проводили в лабораторных условиях под бинокулярным микроскопом.
Было изучено по 30 модельных деревьев на каждом участке, причем на первом провели повторный отбор некоторых тонких моделей, так как их возраст оказался меньше на 3-4 года и, вероятно, это были растения, высаженные в порядке дополнения культур.
Анализ полученных результатов показал, что на участке 1 на шейке корня одна модель имела 73 слоя древесины, четыре – 74 слоя, девятнадцать – 75 слоев и шесть моделей имели по 76 годичных слоев древесины (табл. 6). Среднее число слоев на участке 1 составило на шейке корня 75,0, а на высоте 30 см – 71,4. То есть деревьям понадобилось в среднем 3,6 года для роста в высоту с 5 до 30 см. На участке 2 эту же высоту деревья преодолели всего лишь за 1,9 года. Налицо послепосадочная депрессия роста на первом участке, где культуры были созданы посадкой. По данной таблице мы установили довольно точно биологический возраст культур: 78 лет на первом и 70 лет на втором участке. Этот возраст получается, если считать высоту спила на шейке корня в среднем на высоте третьего годичного прироста растения в высоту.
Таблица 6– Спилы у модельных деревьев в культурах ели
| Участок 1 | Участок 2 | ||||
| Число годичных слоев древесины | Количество спилов, шт. | Число годичных слоев древесины | Количество спилов, шт. | ||
| на шейке корня (5 см) | на высоте пня (30 см) | на шейке корня (5 см) | на высоте пня (30 см) | ||
| – | – | ||||
| – | – | ||||
| – | |||||
| I | – | ||||
| I | – | ||||
| – | I | – | |||
| – | |||||
| Итого: | |||||
| Среднее число годичных слоев | 75,0 | 71,4 | 67,1 | 65,2 |
На нижних спилах и на спиле на высоте груди измерялись диаметры в момент измерения и 10, 20, 30, 40, 50, 55, 60, 65-71 год назад. Соответствие размеров растений в раннем и старшем возрасте мы изучали путем сопоставления диаметров стволика растения в ранние годы с объемом ствола в 70-78 лет. У некоторых моделей удалось «добраться» даже до диаметра стволика в 4 года, однако полные статистические ряды начинались с диаметра в 7 лет на участке 1 и с 5 лет – на участке 2. По фактическим данным были рассчитаны коэффициенты корреляции между размерами растений в раннем возрасте и объемами стволов моделей в 78 лет на участке 1 и в 70 лет – на участке 2. Корреляции оказались весьма близки по значениям, в особенности в самом раннем возрасте (в 7-12 лет) и плавно повышались по параболической кривой, со средними значениями r = 0,32 в 5-6 лет, r = 0,50 в 7-10 лет, r = 0,60 в 11-13 лет и далее с несколько меньшими значениями r в культурах, созданных посевом семян (рис.8). Однако видимые различия оказались недостоверны, хотя они и предполагались до начала исследований.
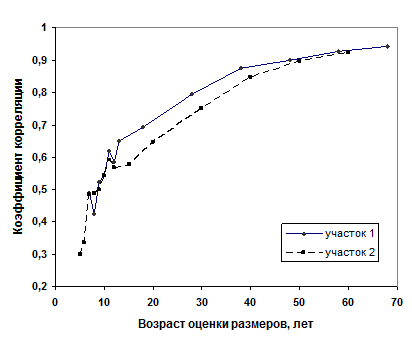
Рис.8 – Соответствие размеров растений в раннем и старшем возрасте на модельных деревьях, отобранных в культурах, созданных посадкой в 1913 г. (участок 1) и созданных посевом в 1919 г. (участок 2).
Затем по всем ранним годам измерения диаметров на спилах рассчитывались средние значения, которые принимали за 100% и все измерения соответствующего года выражали в процентах от среднего. Преимущество таких относительных величин перед абсолютными в том, что данные разных выборок можно объединить в одну совокупность, если они значимо не отличаются одна от другой по некоторым статистическим показателям, например, по коэффициенту изменчивости. Различия по этому показателю между участками 1 и 2 оказались недостоверными для t0,05 во все годы измерений диаметров на спилах и для дальнейшего анализа мы объединили эти выборки и получили совокупность из 60 моделей. Для 5-летнего возраста было получено поле корреляции из 30 точек (на участке 1 статистические ряды для этого возраста были неполными), а начиная с 7-летнего – поля из 60 точек.
Связь между размерами стволов характеризует коэффициент корреляции, который закономерно увеличивался от r = 0,30 в 5 лет до r = 0,65-0,70 в 20 лет (см. рис. 4.2). В каком же возрасте можно начинать отбор лучших растений? Прямого ответа на этот вопрос величина коэффициента корреляции не дает.
Для ответа на него необходимо решить, какие же «деревья будущего» нам нужны. Будем считать, что нам желательны растения, которые формируют стволы с объемами выше среднего значения. Тогда можно разделить поля корреляций на две части горизонтальной линией на уровне 100%. Лежащие выше нее точки можно считать «деревьями будущего» (рис.9). Затем возникает следующий вопрос: из каких растений выросла эта лучшая часть древостоя?

Рис. 9 – Соответствие размеров растений в раннем и старшем возрасте у модельных деревьев в культурах ели.
Если разделить модели по диаметру стволиков в ранние годы на три равные группы - тонкие, средние и толстые и проследить, какие деревья из них выросли, то мы узнаем, какая из групп вносит наибольший вклад в формирование лучших деревьев. Для 5 лет в тонкой, средней и толстой группах оказалось соответственно 10, 9 и 11 точек (см. рис. 4.3).
Посмотрим, сколько крупных деревьев сформировалось из этих групп: из 10 тонких выросло 1 дерево (10%); из 9 средних – 5 (56%), из 11 толстых– 7 (64%). Мы намеренно упростили ситуацию для взрослых деревьев, разбив их на две группы (два ранга). Можно разбить их и на три ранга, однако это усложнит анализ и, в конечном счете, будет давать ответ на тот же вопрос: из чего выросли деревья с желательными для нас размерами. Кроме того, проводимый анализ известен в биометрии как «анализ долей» и для него необходимы два условия. Во-первых, совокупность по результативному признаку делится только на две части: лучшую (желательную для нас) и худшую (нежелательную). Во-вторых, в градациях совокупности (в нашем случае это 3 вертикальных сектора) должно быть не менее чем по 10 наблюдений для доказательства различия на 30% между ними.
Метод анализа долей можно использовать и для прогнозирования. Так, если размеры взрослого дерева соответствуют его размерам в молодости, то это будет оправдавшийся прогноз с надежностью (вероятностью) 100%.
Из данных рисунка следует, что надежность прогноза размеров для тонких растений составит 90% – из 10 таких растений 9 сформировали такие же тонкие стволы (стволы менее среднего объема). Для толстых в 5 лет растений надежность прогноза составит 64% - из 11 растений получилось 7 крупных деревьев. Для средних стволиков прогноз не ясен, так как 56% из них формируют крупные, а 44% – тонкомерные деревья.
При массовом отборе всегда возникает желание повысить его интенсивность и получить более впечатляющие результаты. Это может быть удаление соседей у дерева, выбранного как «самое крупное», с целью еще большего увеличения его прироста. В нашем случае это можно представить как отбор 1/6 части толстых стволиков у моделей и разделение для этого поля корреляции на шесть вертикальных секторов.
Результаты такого увеличения интенсивности отбора показаны на рисунке (рис.10), анализ которого показывает следующее:
- надежность отбора тонких растений при делении на 3 градации составила 75%, а 25% растений сформировали крупные стволы; при делении на 6 градаций надежность отбора увеличилась до 90%, а 10% (1 растение) сформировало крупный ствол;
- надежность отбора толстых растений при делении на 3 градации составила 71% (15 растений из 21 сформировали крупные стволы); при делении на 6 градаций надежность увеличилась до 83% (10 растений из 12 сформировали крупные стволы). 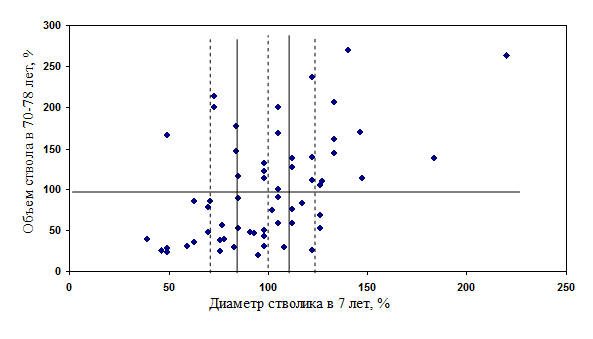

Рис. 10 – Диаметр стволика у ели в 7-летнем возрасте и вероятности формирования крупных модельных деревьев в 70-78 лет при разделении на три и шесть градаций по толщине.
Анализ всех полей корреляции становится более наглядным, если поместить полученные диаграммы для всех возрастов рядом попарно (рис.11). В левой стороне рисунка 11 представлено разделение совокупности на 6 градаций, а в правой – на 3 градации. Из рисунка следует, что только в возрасте 30 лет и старше прогноз роста для тонких и толстых деревьев составляет 100%. Однако необходима ли нам такая точность, если в биометрии уровень безошибочного прогноза обычно принимают 95, а иногда 90, 75 и даже 50%? По-видимому, это зависит от целей работы и задача состоит в выяснении удовлетворяющей нас вероятности такого прогноза.
У наших моделей прогноз роста для 1/3 части толстых стволиков в 5-10 лет составил в среднем 71%, в 12-20 лет – 75% и в 30-40 лет – 90%; для 1/6 части толстых растений эти цифры возрастают до 79, 85 и 100% соответственно.
Здесь в рассуждениях необходимо учесть, что в культурах сохранилось в 1,56-2,73 (в среднем 2,14) раза меньше живых растений по отношению ко всем учтенным – живым и мертвым (см. табл. 5). Можно полагать, что отпад за 70 лет состоял примерно на 80% из тонких и на 20% - из средних по диаметру стволиков. В таком случае принятые нами для выборки из модельных деревьев частоты в градациях (1/3 или 1/6 от количества моделей) в первые годы роста культур были иными и составляли для трех градаций 15,6; 26,2 и 58,2%. То есть соотношение изменилось и доля тонких растений была тогда в 3,74 раза больше. Это можно представить в виде поля корреляции, где левый нижний угол был «заселен» точками гораздо плотнее, чем на наших графиках. Соответственно, положение среднего диаметра стволиков было сдвинуто влево. Поэтому нынешние критерии отбора по размерам для тех лет будут несколько завышены; более правильными будут критерии по интенсивности отбора, учитывающие отпад растений.
| Вероятность формирования крупного дерева в 70-78 лет, % |
| в 40 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| в 30 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| в 20 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| в 12 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| в 10 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| в 7 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| в 5 лет | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Диаметр ствола в раннем возрасте, % от среднего значения |




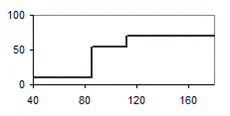







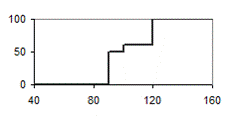

Рис. 11 – Диаметр ели в раннем возрасте и вероятности формирования крупных модельных деревьев в 70-78-летних культурах в условиях густого стояния деревьев, при разделении на три (слева) и шесть (справа) градаций по толщине.
Для корректных расчетов можно принять, что количество тонких в прошлом растений с нынешними размерами диаметра на моделях в интервале от 40 до 70-84% было в 3,74 раза больше и поэтому прогноз формирования из тонких моделей крупных деревьев также понизится в 3,74 раза и составит, например, в 5-7 лет для тонких стволиков не 10-25 %, а не более 6,7%, а для самых тонких не более 2,7%.
Для толстых растений прогноз их роста сохранится, но доля их в первоначальном числе растений в культурах снизится, как и число живых растений, в 2,14 раза. В таком случае при отнесении растения в ранние годы к «толстым» их количество будет не 1/3, а только 15,6% и, соответственно, при отнесении к «самым толстым» не 1/6, а только 7,8%. Примечательна близость этих цифр к доле деревьев 1 класса Крафта в древостое, известная из классического лесоводства (8-15%).
Рассмотренные участки культур в период измерений имели относительную полноту 1,0 и 1,2 и значительное количества валежа и сухостоя говорило о том, что такая полнота сохранялась долгое время, возможно, с 30-летнего возраста, когда густота достигала 2-5 тыс. шт./га. Поэтому почти все тонкие растения в условиях будущей высокой конкуренции уже тогда следовало бы отнести к потенциальному отпаду (примерно 50%) и вырубить их. К возрасту предполагаемой рубки культур мы мало что потеряли бы – подавляющее их большинство сформирует тонкомерные стволы.
Общее представление о надежности отбора растений в раннем возрасте дает рисунок 12. Самое удивительное в результатах проведенного ретроспективного анализа оказалось то, что прогнозы в период от 7 до 20 лет практически не повышаются. Они как бы «сидят» на уровне в 70-80% до 20 лет, что хорошо видно на рисунке.
Такое явление тем более необычно, что несмотря на повышение коэффициентов корреляции с возрастом, соответствующие вероятности не растут. Объяснить это явление можно, если полагать, что в этот период растения никак не могут определиться с лидерами. При этом от лидеров требуется не только скорость роста, мощная крона, но и толерантность к конкуренции. Отсюда следует, что понятия «продуктивность по биомассе» и «конкурентная выносливость» совсем не одно и то же и второе не всегда следует из первого.
Подобные процессы мы исследовали в культурах сосны обыкновенной и получили близкие выводы. Отбор в лидирующую часть древостоя у нее происходит в течение всей жизни, но решающим оказывается период после смыкания крон. Именно в это время в иерархии древостоя происходят наибольшие подвижки. В частности, при повышении густоты в рядах культур корреляции между высотами в 4-7 лет и размерами стволов в 29-40 лет оказались достоверно ниже, вплоть до их отсутствия в отношении высоты (r =0,03-0,19) на одной из пробных площадей (см. табл. 4).
Толерантность к давлению соседей у части деревьев может не сочетаться с продуктивностью по биомассе. По этой причине некоторые особи снижают темпы роста, уступая место не самым продуктивным, но конкурентновыносливым экземплярам. Данное явление показывает, насколько могут быть важны для будущей производительности древостоя опережающие изреживания. Комфортные условия для развития каждого дерева стабилизируют ранговую структуру древостоя, что существенно повышает производительность как отдельных растений, так и древостоя в целом.

Рис.12 – Вероятности формирования крупных в 70-78 лет модельных деревьев из тонких, самых тонких, толстых и самых толстых стволиков в 5-40 лет.
Выводы
Таким образом, изучение роста модельных деревьев созданных в 1913 и 1919 г. по схемам Ф. А. Теплоухова, позволила установить следующее. В 70-78-летних культурах ели с высокой относительной полнотой (1,0-1,2) и густотой, достигавшей 2-5 тыс. шт./га в 20-30 лет, при разделении моделей в раннем возрасте по диаметру стволика на 3 и 6 градаций, можно дать следующие ретроспективные вероятности формирования из них деревьев с объемом ствола выше среднего (крупных деревьев).
1. Для растений с толстыми стволиками (15,6% количества растений в раннем возрасте) ретроспективные вероятности формирования из них крупных деревьев составят: в 5 лет 64%, в 7-12 лет 70-80%, в 20лет 70%, в 30 лет 80% и в 40 лет 100%.
2. Для растений с самыми толстыми стволиками (7,8% количества растений в раннем возрасте) указанные вероятности возрастают: в 5 лет 67%, в 7-12 лет 80-83%, в 20 лет 90%, в 30 лет и далее 100%.
3. Тонкие растения (до 60% количества растений в раннем возрасте) в любом возрасте, начиная с 5-7 лет, имеют мало шансов (менее 7%) сформировать крупные деревья, а самые тонкие (30% всех растений) их почти не имеют (ретроспективная вероятность формирования из них крупного дерева 2,7%).
Заключение о надежностях прогноза роста деревьев ели сибирской в культурах
Ранняя диагностика размеров деревьев у ели сибирской, по отношению к их размерам в 70-78 лет в культурах, возможна при следующих надежностях прогноза:
- «мелкие» деревья (объем ствола ниже среднего) диагностируются в 5-7 лет по диаметру стволика менее 80% от среднего, с надежностью прогноза 93-97%;
- «крупные» деревья (объем ствола выше среднего) диагностируются в 7-10 лет по диаметру стволика 110% и более, с надежностью прогноза 70-75% и его улучшением до 85% при отборе по диаметру 125% и более.
Заключение подтверждает открытый в культурах сосны ранговый закон роста деревьев в древостое Е.Л.Маслакова (1984).
|
из
5.00
|
Обсуждение в статье: Отбор быстрорастущих деревьев ели в возрасте 5, 7, 12, 30, 40 лет и формирование из них крупных деревьев (ретроспекция развития) |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы