 |
Главная |


ОСНОВНЫЕ ПОЛОЖЕНИЯ НЕЙРОННОЙ ТЕОРИИ
|
из
5.00
|
Нервными окончаниями называются концевые разветвления отростков нервных клеток, в которых нервный импульс или генерируется, или передается на другую клетку (структуру).Все нервные окончания по функции делятся на три группы:
1.Эффекторные нервные окончания.
2.Рецепторные, или афферентные, нервные окончания.
3.Межнейронные синапсы.
ЭФФЕКТОРНЫЕ НЕРВНЫЕ ОКОНЧАНИЯ. Их функцией является вызывание эффекта, в зависимости от которого они делятся на две группы: 1) двигательныеи 2) секреторные.Двигательные окончания подразделяются на: 1) двигательные окончания в скелетной мышечной ткании 2) двигательные нервные окончания в гладкой мышечной ткани.
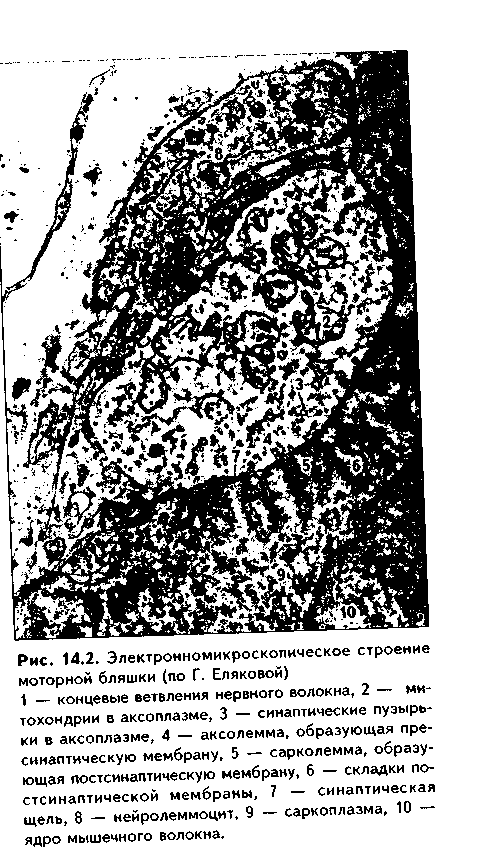
Двигательные нервные окончания в скелетной мышечной ткани называются нервно-мышечным синапсом,или моторной бляшкой(рис. 14.1,14.2). Представляют собой окончания аксонов мотонейронов передних рогов спинного мозга на поперечнополосатых мышечных волокнах. При образовании моторной бляшки миелиновое нервное волокно, подходя к мышечному волокну, теряет миелиновую оболочку. Осевой цилиндр внедряется в мышечное волокно, прогибая сарколемму. Терминальное ветвление аксона имеет на конце утолщение. Это так называемый нервный полюснервно-мышечного синапса. В нем обнаруживаются митохондрии, микротрубочки (нейротрубочки), синаптические пузырьки с нейромедиатором ацетилхолином.Размеры последних составляют около 50 нм. В пресинаптической мембране есть утолщения — активные зоны,где происходит выделение(секреция) медиатора.

|
Плазмолемма мышечного волокна и прилегающая саркоплазма образуют мышечный полюс.Между пресинаптической и постсинаитической мембранами находится синаптическаящель. Она имеет ширину 50—100 нм и содержит базальную мембрану и отростки глиоцитов. На базалыюй мембране имеются сигнальные белки агрин, S-ламинини др., которые служат мет- ками, при помощи которых регенерирующий аксон мотонейронов находит синапти-ческую зону на мышечном волокне.
Постсинаптическая мембрана имеет много складок, которые образуют вторичные синаптические щели.Они во много раз увеличивают поверхность синаптической щели и содержат материал базальной мембраны. В по-стсинаптической мембране имеются никотиновые холи-норецепторы,концентрация которых достигает 20—30 тыс. на 1 мкм2. В зоне синапса мышечное волокно теряет исчерченность (мио-фибриллы лежат глубже зоны синапса), но содержит большое количество митохондрий, профилей гранулярной ЭПС, рибосом, скопление ядер.
МЕХАНИЗМ РАБОТЫ НЕРВНО-МЫШЕЧНОГО СИНАПСА (см. рис. 12.10). Нервный импульс доходит до пресинантического полюса и вызывает увеличение мембранной проницаемости этого полюса для ионов кальция. При этом концентрация кальция в пресинаптическом полюсе резко возрастает благодаря: 1) высвооождению его из депо (гладкой ЭПС, митохондрий), а также 2) поступлению из внеклеточной среды. Далее кальций вызывает взаимодействие компонентов цитоске-лета, которые содержатся в пресинаптическом полюсе. Очевидно, наиболее выражены кинезиновыйи динеиновыймеханизмы транспорта синаптичес-ких пузырьков к пресииаптической мембране. После перемещения синаптических пузырьков к пресииаптической мембране происходит слияние с ней мембран, окружающих пузырьки, а затем пузырьки раскрываются в синаитическую щель и < выделяют в нее медиатор. Далее медиатор мигрирует к иостсинаптичес-кой мембране и вызывает ее деполяризацию, сливаясь с рецепторами ацетилхолина.
Деполяризация плазмолеммы мышечного волокна передается по Т-трубочкам на всю толщину мышечного волокна, а затем с Т-тру-бочек переходит на терминальные цистерны сарконлазматического ретикулума (СПР). Это вызывает увеличение проницаемости СПР для ионов Са2+, который выходит из СПР и мигрирует к актиновым филаментам. Там он вызывает конформационные изменения в молекуле тропонина и открывает активные центры на ак-
активные центры >ш сатиновых филаментах. С этими центрами начинают связываться головки миозина. Происходит мышечное сокращение.
В иостсинаптической мембране содержится фермент ацетилхолшюсте-раза,который разрушает избыток ацетилхолина в синаптической щели и уменьшает время действия медиатора. Это необходимо для предотвращения перевозбуждения иостсинаптической мембраны.
Патология нервно-мышечного синапса.При отравлении фосфорорга-ническими соединениями (ФОС), которые относятся к боевым отравляю- щим веществам и широко используются в быту как инсектициды, актин ность ацетилхолинэстеразы подавляется. При этом в синаптической щели накапливается медиатор, вызывающий перевозбуждение иостсинаптичес-кой мембраны. В результате возникают судорожные сокращения мышц, затем сменяющиеся параличом. От паралича межреберных мышц больной погибает. Для лечения отравлений ФОС применяют реактиваторы ацетилхолинэстеразы,которые восстанавливают активность холинэстеразы и работу нервно-мышечного синапса.
Блокада ацетилхолиновых рецепторов на постсинаптической мембране может быть осуществлена некоторыми ядами (яд кураре).При этом становится невозможной передача возбуждения с нервного окончания на мышцу, и мышца полностью расслабляется. Синтетические аналоги кураре (курареподобные вещества, миорелаксанты)используются в хирургической практике для расслабления мышц при операциях. Существует заболевание миастения гравис,при котором в результате аутоиммунной реакции происходит разрушение ацетилхолиновых рецепторов в постсинаптической мембране нервно-мышечного синапса. Характеризуется прогрессирующей мышечной слабостью.
Двигательные нервные окончания на гладких мышцахпредставляют собой варикозные расширения терминалей аксона, которые контактируют с одним из миоцитов в миоцитарном комплексе. Терминали содержат си-наптические пузырьки с ацстилхолином или норадреналином.
Секреторные нервные окончанияпредставляют собой терминали аксонов, которые вступают в тесную связь с секреторными клетками: или подходят к ним, не проникая через базальную мембрану, или пенетрируют ба-зальиую мембрану и вдавливаются в секреторные клетки, образуя терминальные расширения. Нейролемма аксона и плазмолемма секреторной клетки образуют соответственно пре- и постсинаптические мембраны, разделенные узкой синаптической щелью. Медиатор, выделившийся из синаптичес-ких пузырьков, вызывает деполяризацию мембраны секреторной клетки, что приводит к высвобождению кальция из депо (обычно он находится в митохондриях и секреторных гранулах). Кальций связывается с белком кальмо-дулином,и этот комплекс вызывает два эффекта: полимеризацию микротрубочек и взаимодействие актиновых и миозиновых филамент, что способствует продвижению секреторных пузырьков к цитолемме, слиянию их мембраны с цитолеммой и ведет к последующему выделению секрета из клетки.
|
из
5.00
|
Обсуждение в статье: ОСНОВНЫЕ ПОЛОЖЕНИЯ НЕЙРОННОЙ ТЕОРИИ |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы