 |
Главная |


Перенос ионов через каналы биомембран
|
из
5.00
|
Этот вид переноса характерен в первую очередь для возбудимых (нервных и мышечных) клеток. Каналы обычно предназначены для создания интенсивных потоков определенных видов ионов, если на мембране существует градиент их концентрации. Именно поэтому каждому виду канала должен соответствовать ионный насос, создающий этот градиент (см. раздел 1.5. - активный транспорт).
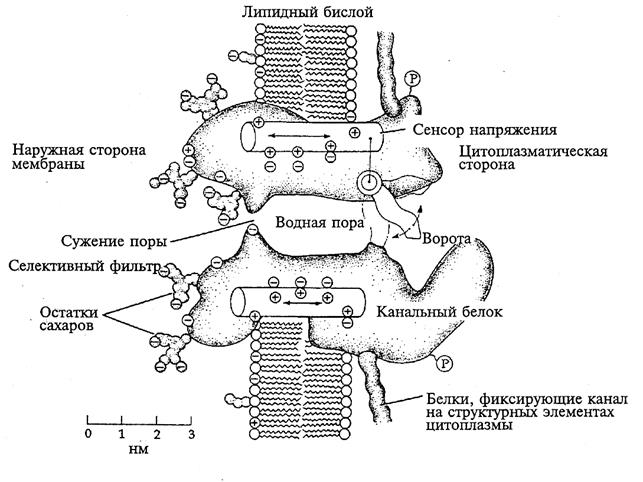 |
Ионные каналы (рис.5) - это мембранные структуры, представляющие собой интегральные белки (гликопротеины).
Рис.5. Схематическое изображение ионного канала (по B. Hille, 1992).
Канал изображен в виде расположенной в мембране макромолекулы со сквозной полостью посередине. Часть молекулы, обращенная в наружный раствор, несет гликозидные группы. Представления о функциональных участках канала - селективном фильтре, воротах и сенсоре напряжения - основаны преимущественно на методе фиксации напряжения и дополнены данными структурных исследований.
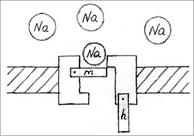
Ионные каналы пронизывают липидный слой и способны при адекватных внешних воздействиях (изменение потенциала на мембране, действие гормона или медиатора) избирательно менять проницаемость мембраны для определенных ионов (Na+, K+, Ca+2, Cl-). Например, (рис.6) в покое (мембрана не деполяризована, см. раздел 2.3). Натриевый канал не пропускает ионы Na+, поскольку закрыты 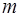 -ворота ( - и
-ворота ( - и 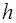 -ворота -
-ворота - 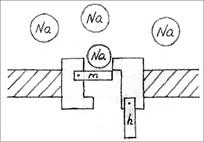
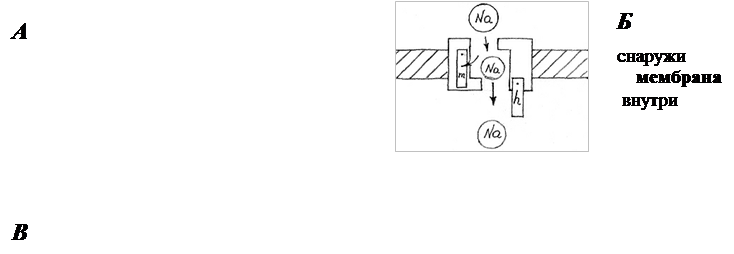
это потенциалзависимые субъединицы белка-канала).

|

 | |||
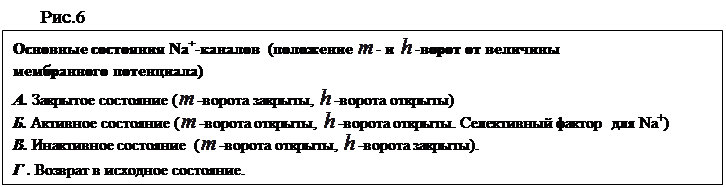 |
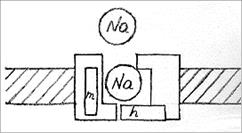
При деполяризации -ворота открываются, и канал активируется, т.е. начинает пропускать ионы Na+. Из-за этого -ворота называют также активационными. В открытом состоянии проводимость канала в значительной степени определяется его селективным фильтром, который не пропускает анионы и гораздо более свободно пропускает Na+, чем K+ или Ca+2. При более длительной деполяризации закрываются -ворота (инактивационные), расположенные у внутренней стороны мембраны, и канал инактивируется. Реполяризация до уровня потенциала покоя вновь приводит к открытию
-ворот и закрытию -ворот. В этом состоянии канал можно вновь активировать деполяризующим стимулом (см. раздел 2.4.)
В основе многих физиологических процессов (передача электрических и химических сигналов, мышечное сокращение, секреторный процесс и т.д. ) лежит прежде всего работа ионных каналов. Их характеристики могут изменять некоторые фармакологические препараты и яды. Существуют блокаторы ионных каналов, например, лекарственные вещества, антагонисты кальция (верапамил, нифедипин и др.) они временно блокируют ионные каналы, снижая мышечный тонус сосудов. Есть лекарственные вещества, временно блокирующие натриевые каналы, например, анестетик - лидокаин, новокаин. Они снижают местную чувствительность, устраняют чувство боли. Необратимо блокируют ионные каналы такие паралитические яды, как тетродоксин, вырабатываемый рыбой Spheroidus rubrides (фугу) или сакситоксин, продуцируемый некоторыми планктонами. Сильный блокирующий эффект калиевых ионных каналов оказывает тетраэтиламмоний. Существуют также активаторы, открывающие ионные каналы, например, лекарственный препарат миноксидил, активирует калиевые каналы.
Каналам биомембран свойственна характерная избирательность для ионов (селективность), а также способность открываться и закрываться при различных воздействиях на мембрану (воротная функция). Переходы каналов из открытого состояния в закрытое (воротный механизм) могут быть обусловлены изменениями мембранного потенциала, взаимодействием с определенными химическими веществами, специфическим фосфорилированием каналов. Воротный механизм каналов управляется сенсором внешнего стимула. В зависимости от локализации сенсора каналы разделяются на две группы.

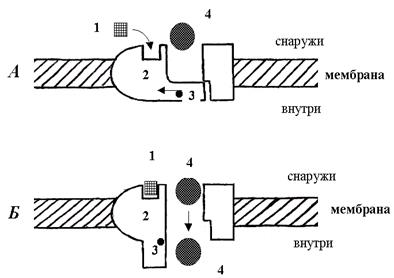 К первой группе относятся каналы (рис. 7), имеющие собственный сенсор (входящий в состав макромолекулы) внешнего сигнала.
К первой группе относятся каналы (рис. 7), имеющие собственный сенсор (входящий в состав макромолекулы) внешнего сигнала.
Рис. 7. Схема работы ионных каналов I типа, имеющего собственный сенсор
для внешнего сигнала.
Внешний стимул влияет непосредственно на макромолекулу канала. Эта группа включает два больших семейства ионных каналов: потенциал - и лигандзависимые.
Потенциалзависимые каналы (Na+, K+, Ca+2-каналы) открываются и закрываются при изменении электрического потенциала на мембране.
Лигандзависимые ионные каналы обеспечивают быструю передачу сигналов между клетками, например, в химических синапсах. Эти каналы открываются при связывании с рецептором ряда биологически активных веществ, таких как ацетилхолин, глутамат,
-аминомасляная кислота.
|
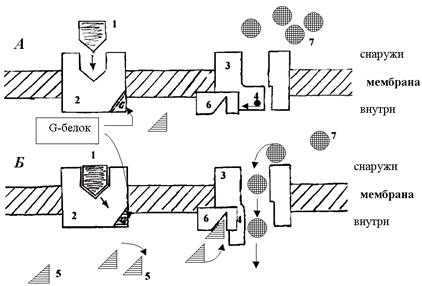 |
В каналах второй группы (рис.8) сенсор внешнего сигнала (рецептор первичного посредника) пространственно разобщен с каналом.
Рис.8. Схема работы ионного канала II типа, имеющего внешний сенсор от него, и поэтому канал опосредованно управляется вторичным посредником (химическим сигналом).
Взаимодействие сенсора и канала осуществляется с помощью растворимых внутриклеточных вторичных посредников. Это рецепторзависимые ионные каналы, каналы, опосредованно управляемые химическими сигналами. К ним относятся также каналы, управляемые G -белками, которые активируются при связывании лиганда с рецептором.
Кроме вышеописанных каналов в мембране находятся неспецифические каналы для пассивного транспорта ионов (в первую очередь для ионов калия). Эти каналы не имеют воротных механизмов, всегда открыты и почти не меняют свое состояние при электрических воздействиях на мембрану.
Осмос
Клеточные мембраны обладают свойством полупроницаемости, то есть способностью хорошо пропускать одни вещества, например, воду, и плохо другие. Вода проходит в результате осмоса. Осмос- это движение молекулы воды через полупроницаемую мембрану из области меньшей в область большей концентрации растворенного вещества. Сила вызывающая это движение - осмотическое давление. Осмос, по существу, представляет собой диффузию молекул растворителя. За меру осмотического давления принимают то механическое давление, например гидростатическое, которое уравновешивает осмотическое давление и выравнивает потоки молекул растворителя в одну и другую стороны. Осмотическое давление зависит от количества растворенных частиц и от температуры.  P=iRCT R- универсальная газовая постоянная,
P=iRCT R- универсальная газовая постоянная,
i - изотонический коэффициент - показывает во сколько раз увеличивается количество частиц при диссоциации молекулы вещества. Для неэлектролитов i=1.
Осм. давление крови человека 7,6–7,8 атм.
Фильтрация
Кроме осмоса, перенос воды может осуществляться путем фильтрации при наличии градиента гидростатического давления. Фильтрация - движение жидкости через поры какой-либо перегородки под действием гидростатического давления. Скорость фильтрации dV/dt находят из уравнения Пуазейля, описывающего истечение жидкости из капилляра под давлением.
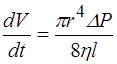
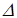 Р - разность давлений в начале и в конце поры, r- радиус поры,
Р - разность давлений в начале и в конце поры, r- радиус поры,
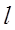 - длина поры,
- длина поры, 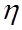 - коэффициент вязкости жидкости.
- коэффициент вязкости жидкости.
1.4.4.Водный обмен между кровью и лимфой

Явление фильтрации и осмоса имеют особое значение в процессе обмена воды между кровью и тканями. Осмотическое давление крови человека 7,6 - 7,8 атмосфер. Это давление является суммой давлений всех растворенных в плазме крови веществ.
Особое значение в водном обмене между кровью и тканевой жидкостью имеет та часть общего осмотического давления, которая обусловлена высокомолекулярными веществами - белками. Эта часть осмотического давления называется онкотическим давлением. Величина осмотического давления в 200 раз превосходит величину онкотического давления. Несмотря на это онкотическому давлению принадлежит основная роль в поступлении воды в кровеносное русло из тканевой жидкости.
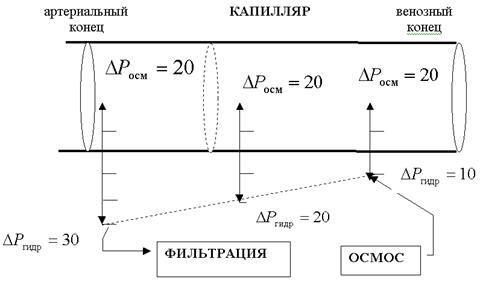
Онкотичесое давление крови человека равно 30 мм.рт.ст. ,а тканевой жидкости и лимфы 10 мм.рт.ст. Под действием разности онкотического давления крови и лимфы величиной 20 мм.рт.ст. вода поступает из лимфы в кровь. Одновременно с градиентом онкотического давления ( 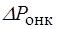 ) существует и градиент гидростатического давления,
) существует и градиент гидростатического давления,
обусловленный работой сердца ( 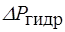 ). В артериальном конце капилляра гидростатическое давление крови 30 мм.рт.ст.. На протяжении капилляра кровяное давление падает: в середине оно равно 20 мм.рт.ст, а в венозном конце - 10 мм.рт.ст..
). В артериальном конце капилляра гидростатическое давление крови 30 мм.рт.ст.. На протяжении капилляра кровяное давление падает: в середине оно равно 20 мм.рт.ст, а в венозном конце - 10 мм.рт.ст..
 | |||
| |||
Соответственно, если сравнить значения градиентов онкотического и гидростатического давлений в различных участках капилляров, то очевидно, что в артериальном конце преобладает гидростатическое давление - наблюдается фильтрация, в результате которой вода выходит из кровяного русла, а в венозном конце преобладает онкотическое давление, т.е. наблюдается осмос, и вода поступает из ткани в лимфу (рис.9).
Цифрами обозначены осмотическое и гидростатическое давления в мм.рт.ст.
В норме процессы фильтрации и осмоса скомпенсированы. При патологиях, например: лучевая болезнь – нарушение проницаемости мембран, при долгих голоданиях, гипертонии, шоке, ожогах, при больших кровопотерях уменьшается градиент онкотического давления, и тогда фильтрация воды преобладает над осмосом, вследствие этого наблюдаются отеки.
Активный транспорт
Перенос молекул и ионов против электрохимического градиента, осуществляемый клеткой за счет энергии метаболических процессов, называется активным транспортом.
Осуществляя такой транспорт, клетка совершает работу, которая называется концентрационной или осмотической. Накопление клеткой вещества, содержащегося в окружающей среде в малых количествах, и выведение тех веществ, концентрация которых в окружающей среде значительно больше, чем в клетке, обеспечивается активным транспортом.
A=mRTln 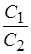 A-концентрационная работа, m-количество молей вещества, перенесенных через мембрану, T -абсолютная температура,
A-концентрационная работа, m-количество молей вещества, перенесенных через мембрану, T -абсолютная температура, 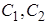 -концентрации ионов внутри и вне клетки.
-концентрации ионов внутри и вне клетки.
У человека в покое примерно 30 - 40 % всей энергии, образующейся в ходе метаболических процессов, расходуется на активный транспорт. В тканях, где активный транспорт особенно интенсивен, потребляется много кислорода даже в покое; так мозг человека составляет 1/50 массы тела, а потребляет в покое 1/5 всего кислорода. Активный транспорт - одно из удивительных свойств живых организмов.
Активный транспорт веществ через биологические мембраны имеет огромное значение. За счет активного транспорта в организме создаются градиенты концентраций, градиенты электрических потенциалов, градиенты давления и т.д., поддерживающие жизненные процессы, то есть с точки зрения термодинамики активный перенос удерживает организм в неравновесном состоянии, поддерживает жизнь.
Существование активного транспорта веществ через биологические мембраны впервые было доказано в опытах Уссинга (1949г.) на примере переноса ионов натрия через кожу лягушки. Экспериментальные данные неопровержимо свидетельствовали о том, что перенос ионов натрия через кожу лягушки не подчиняется уравнению пассивного транспорта. Следовательно, имеет место активный перенос.
Изучение систем активного транспорта, называемых биологическими насосами, является важной задачей исследования клеточных мембран. Нарушение активного транспорта приводит к гибели клетки. Отключение Nа+- К+ насоса на 10 минут приводит к изменению ионного состава внутри клетки на 40 - 50 %.
В покое мембрана не является абсолютно непроницаемой для любого вида ионов. Диффузионные потоки, не будь им противодействия, рано или поздно выровняли бы состав по обе стороны мембраны. Для восстановления и поддержания необходимой концентрации ионов внутри клетки в мембране содержится специальный аппарат, обеспечивающий их активный транспорт против градиента их собственной концентрации.
Активный транспорт ионов Nа+, К+ и Са+ осуществляется двумя типами "насосов"
Nа+ - К+ и Nа+ - Са2+.
Наиболее характерная черта таких насосов - перенос вещества против градиентов. Следовательно, необходимо энергетическое обеспечение такого процесса. Организм должен за счет своей свободной энергии преодолеть физико-химические градиенты, повернуть вспять движение веществ. Значит, второй особенностью активного транспорта является наличие источника энергии для его осуществления. Причем эта энергия поступает из химических реакций. Рассмотрим общие принципы превращения энергии в организме.
Клетки получают свободную энергию за счет освобождения энергии химических связей, заключенной в питательных веществах. Они, в свою очередь, получают эту энергию от Солнца. На земле солнечную энергию улавливают зеленые растения, их пигмент хлорофилл - поглощает энергию солнечного света. Солнечная энергия переводит электроны в молекулах хлорофилла с основного энергетического уровня на более высокий. Возбужденные электроны стремятся возвратиться на основной (устойчивый) уровень. Возврат сопровождается отдачей энергии. В клетках хлорофилл связан с другими специфическими молекулами (переносчиками), поэтому возбужденные электроны, отрываясь от молекул хлорофилла, переносятся по цепи реакций, в которых участвует переносчик. Энергия, отданная электронами промежуточным веществам, используется на образование глюкозы, из которой в свою очередь могут образовываться аминокислоты и жиры. В процессе их созидания солнечная энергия запасается в химических связях. Так, например, одна грамм-молекула глюкозы несет 690 ккал. Организм человека и животных извлекает эту энергию при окислении питательных веществ: белков, углеводов, жиров. Такой процесс называется тканевым дыханием. Вне организма (окисление, горение). При тканевом дыхании происходит постепенное окисление.
Энергия, синтезированная в химическую форму, удобна для транспортировки, во-первых, к клеткам и внутрь клетки по всем органеллам. Во-вторых, форма "консервирования" свободной энергии должна быть такой, чтобы она позволяла клетке легко пользоваться этой энергией, таким требованиям удовлетворяют макроэргические соединения, в частности аденозинтрифосфат (имеет три фосфатные группы). Энергия из АТФ извлекается при гидролитическом расщеплении этой молекулы, при этом каждая грамм моль АТФ поставляет 7-8 ккал. и превращается в АДФ. С позиции термодинамики АТФ - заряженная форма носителя энергии, а в АДФ - разряженная. Переход АДФ в АТФ происходит в митохондриях при окислении питательных веществ - зарядка. В клетке в процессе гидролиза АТФ кольцевая фосфатная группа переносится на молекулы другого вещества, которые при этом фосфорилируются, то есть присоединяют фосфатную
группу, приобретая свободную энергию. Такая "возбужденная" молекула способна участвовать в процессах, нуждающихся в поставке энергии, в том числе и в активном транспорте. Таким образом, система активного транспорта (биологические насосы) включает источник свободной энергии и связанный с ним посредством химических реакций переносчик транспортируемого вещества. Механизм может быть различен. Это либо малые белковые молекулы, входящие в состав мембран, проходят через всю мембрану по типу карусели, либо крупные белковые молекулы, насквозь пронизывающие мембраны, приняв молекулу переносимого вещества и фосфатную группу, совершают поворот на 1800 и отдают транспортируемые вещества в примембранное пространство с противоположной стороны, затем занимают исходное положение, т.е. ротационный механизм.
Процесс гидролиза АТФ в пробирке течет крайне медленно, для его активации надо добавить ферменты АТФ-азы, содержащиеся в мембранах, следовательно, кроме макроэргов (АТФ) переносчиков есть третий элемент, обеспечивающий сопряжение в работе первых двух. Для активного транспорта каждого вещества (каждого иона) обнаружена своя АТФ-аза. Вся система включается тогда, когда концентрация транспортируемого вещества становится выше какого-то предела. Это вещество активирует специфическую АТФ-азу. В результате осуществляется гидролиз АТФ, отщепившаяся фосфатная группа фосфорилирует переносчик. В результате этого переносчик приобретает энергию достаточную для трансмембранного переноса веществ против концентрационного градиента.
Рассмотрим этот процесс на примере К+-Nа насоса. Ионы К+ и Nа+ вводятся и выводятся из клетки и в клетку против концентрационного градиента. Клетка очень строго следит за содержанием в своей цитоплазме названных ионов (рис.10).
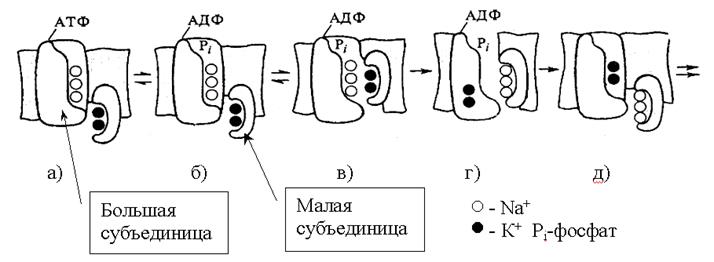
Рис.10. Схема рабочего цикла Na-K насоса. Рабочий цикл димера Na+ К+ - насоса.
а) связывание Na+ и К+ в полостях субъединиц,
б) Na+ -зависимое фосфорилирование большой субъединицы,
в), г) обмен катионами между субъединицами,
д) K+-зависимое дефосфорилирование и выброс ионов.
При повышении концентрации ионов натрия активируется так называемая калий-натрий зависимая АТФ-аза, этот фермент содержащийся в плазматической мембране, активирует гидролиз АТФ.
Активируя гидролиз АТФ, он обеспечивает фосфорилирование переносчика, который выносит натрий из клетки. На внешней поверхности мембраны происходит ионный обмен натрия на калий внутри ферментного комплекса, после этого обмена белок-переносчик совершает обратный переворот с переносом калия внутрь клетки. Ион калия и неорганическая фосфатная группа (Р) освобождается от белка-переносчика, который в свою очередь возвращается в исходное состояние. Подсчитано, что за полный цикл 1 молекула АТФ обеспечивает перенос через клеточную мембрану 3 иона Na+ и
2 иона К+.
К+ – Na+ насос работает очень напряженно, особенно в нервных клетках. При этом из клетки уносится один положительный заряд. Следовательно, Na+ – К+ насос электрогенный, т.е. он участвует в создании потенциала покоя на мембране, увеличивая его примерно на 25% (см. раздел 2.3.), хотя это и не основной механизм в создании потенциала покоя (основной механизм связан с полупроницаемостью мембраны), но это основной механизм поддержания градиентов концентрации Na+-K+ на мембране. Существуют лекарственные вещества - сердечные гликозиды (дигоксин, уабаин, строфансин, коргликон и т.д.), которые ингибируют Na+-K+ АТФ-азу. Эти вещества обладают токсической активностью.
В клеточной мембране постоянно присутствуют и переносчики, и транспортные АТФ-азы, в примембранном пространстве клетки находится АТФ, выходящий из митохондрий, которые подтягивают к местам активного транспорта. Однако, вся система не работает до появления определенного стимула, которым обычно служит нарастание концентрации вещества, подлежащего активному транспорту. Этот фактор активирует специфическую АТФ-азу, которая в свою очередь катализирует гидролиз АТФ с отщеплением концевой фосфатной группы. Присоединяясь к переносчику, она фосфорилирует его. При фосфорилировании переносчик приобретает дополнительную свободную энергию, необходимую и достаточную для трансмембранного переноса вещества вопреки действию физико-химических градиентов. Клеточные мембраны проявляют весьма высокую селективность по отношению ко многим веществам, причем, разные клетки берут из межклеточной среды обычно то, что им требуется. Наборы биологических насосов в мембранах клеток разных типов неодинаковы. Интенсивность их функционирования так же различна. Многие клетки имеют, например, системы активного транспорта глюкозы ("сахарные насосы"), но ни одна из них не может сравниться с нервной по интенсивности транспорта этого вещества. Особенно высока интенсивность клеток к аминокислотам-тем "кирпичикам", из которых строятся белки, следовательно, построение белковых молекул, обеспечивающих основные процессы жизнедеятельности в клетках, тесно связано с транспортом аминокислот через мембраны. У человека весом 70 кг ежечасно обновляется около 100 граммов белка.
|
из
5.00
|
Обсуждение в статье: Перенос ионов через каналы биомембран |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы