 |
Главная |


Превращение глюкозо-6-фосфата в триозофосфаты
|
из
5.00
|
УГЛЕВОДЫ
Углеводы — органические вещества, сод карбонильную группу и несколько гидроксильных групп. По способности к гидролизу на мономеры углеводы делятся на две группы: простые (моносахариды) и сложные (дисахариды и полисахариды). Биологическая роль-Структурная и опорная функции. Углеводы участвуют в построении различных опорных структур. Так целлюлоза является основным структурным компонентом клеточных стенок.Защитная роль у растений.Пластическая функция. Углеводы входят в состав сложных молекул участвуют в построении АТФ, ДНК и РНК).Энергетическая функция. Углеводы служат источником энергии: при окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды.Запасающая функция. Углеводы выступают в качестве запасных питательных веществ: гликоген, крахмал и инулин.Осмотическая функция. Углеводы участвуют в регуляции осмотического давления в организме. Так, в крови содержится 100—110 мг/% глюкозы, от концентрации глюкозы зависит осмотическое давление крови.Рецепторная функция. Олигосахариды входят в состав воспринимающей части многих клеточных рецепторов или молекул-лигандов.
Расщепление углеводов начинается в полости рта и продолжается в желудке. Катализатором гидролиза является фермент α‑амилаза слюны. При расщеплении из крахмала и гликогена образуются декстрины и в небольшом количестве - мальтоза. Пережеванная и смешанная со слюной пища в желудок. Проглоченные пищевые массы перемешиваются с желудочным соком, содержащим соляную кислоту. Желудочный сок не содержит ферментов, расщепляющих сложные углеводы. Поэтому гидролиз углеводов с увеличением кислотности в желудке прерывается и возобновляется в двенадцатиперстной кишке. В двенадцатиперстной кишке происходит наиболее интенсивное переваривание крахмала и гликогена с участием α‑амилазы сока поджелудочной железы. В двенадцатиперстной кишке кислотность значительно снижается. Среда становится практически нейтральной, оптимальной для максимальной активности α‑амилазы панкреатического сока. Поэтому гидролиз крахмала и гликогена с образованием мальтозы, в тонкой кишке завершается. Процессу гидролиза с участием α‑амилазы панкреатического сока дополнительно способствуют еще два фермента: амило-1,6-глюкозидаза и олиго-1,6-глюкозидаза (терминальная декстриназа).
Углеводы всасываются в виде моносахаридов. Активнее всасываются глюкоза и галактоза. их всасывание обеспечивается тесно связанным с Na + трансмембранным транспортом. В апикальной мембране содержится особый белок - переносчик Na + и глюкозы. Он имеет два места - одно «садится» Na +, на второе - глюкоза. На внутренней поверхности мембраны переносчик освобождается от Na + и глюкозы и возвращается обратно. Это вторичный активный транспорт глюкозы. Na + сначала по градиенту концентрации достигает базолатерального мембраны, а затем откачивается насосом. Глюкоза переходит через безолатеральни мембраны с концентрационным градиентом. В случае отсутствия Na + или при блокаде насоса глюкоза всасывается в 100 раз медленнее. При этом, вероятно, «работает» механизм простой диффузии. Путем простой диффузии в клетки поступает манноза, а путем облегченной - фруктоза. скорость всасывания глюкозы неодинакова. В тощей кишке в 3 раза выше, чем в подвздошной. Некоторые аминокислоты тормозят всасывание глюкозы, а глюкокортикоиды, тироксин, инсулин, серотонин усиливают. Гистамин и соматостатин тормозят. Парасимпатические нервы стимулируют, а симпатичные тормозят этот процесс.превращение в усваиваемую организмом глюкозу, зависит от их гликемического индекса. Гликемический индекс углевода определяет способность углевода повышать гликемию, то есть количество глюкозы в крови. Другими словами, ГИ выражает способность углевода к гидролизу, то есть расщеплению до глюкозы. Если ГИ равен 100, это значит, что при попадании в тонкую кишку она всосется через стенки кишечника на 100 %. Если ГИ белого хлеба равен 70, это означает, что содержащийся в нём углевод (крахмал) на 70% гидролизуется и пройдет через стенки кишечника в форме глюкозы.
По характеру действия на полисахариды различают α-, β- и γ-амилазы: под влиянием α-амилаз полисахариды гидролизуются до олигосахаридов, β-амилазы отщепляют от внешних цепей молекулы полисахарида дисахарид мальтозу, γ-амилазы — моносахарид глюкозу. Определение активности α-амилазы в сыворотке крови и моче используется в диагностике ряда заболеваний в качестве дополнительного диагностического теста. Наиболее информативно и достоверно повышение активности А. в крови при заболеваниях поджелудочной железы, происходящее за счет увеличения активности панкреатического изофермента α-амилазы. Активность А. в крови и моче резко возрастает при остром панкреатите и раке поджелудочной железы. Увеличение активности А. в крови отмечают также при вирусном гепатите, поражении слюнных желез, почечной недостаточности, лечении кортикостероидами, салицилатами, антибиотиками тетрациклинового ряда, фуросемидом, гистамином. Активность α-амилазы в биологических жидкостях определяют по количеству гидролизованного ею растворимого крахмала (амилокластический метод). В качестве унифицированных в СССР утверждены два амилокластических метода: метод Каравея и метод Смита — Роу. Метод Каравея основан на том, что α-амилаза расщепляет крахмал на продукты, не дающие цветной реакции с йодом; по уменьшению интенсивности окраски судят об активности фермента. Нормальные величины активности α-амилазы, определенные методом Каравея, составляют: в сыворотке крови 3,3—8,9 мг/(с ․л), или 12—32 мг/(ч․мл), в моче до 44 мг/(с․л), или до 120 мг/(ч․мл), в дуоденальном содержимом 1,7—4,4 г/(с․л), или 6—16 г/(ч․мл). Амилокластический метод определения активности α-амилазы по Смиту — Роу основан на фотометрическом измерении уменьшения концентрации растворимого крахмала в результате его гидролиза амилазой. Нормальные величины активности α-амилазы, определенной по методу Смита — Роу, составляют в крови — 16—30 мг, в моче — до 160 мг крахмала, гидролизованного амилазой, содержащейся в 1 мл крови или мочи, за 1 ч инкубации при 37°.
Пути тканевых превращений
Глюкоза и гликоген в клетках распадаются анаэробным и аэробным путями. Аэробный распад характерен для тканей, обогащенных кислородом, анаэробный – для тканей с недостатком О2. Анаэробный распад глюкозы происходит при недостаточном содержании кислорода, в клетках мышечной ткани животного организма. Данный путь распада называется дихотомическим, т.к. в процессе происходит образование двух молекул триоз, содержащих по 3 С-атома из одной молекулы гексозы (6 С-атомов). Конечный продукт анаэробного превращения глюкозы – молочная кислота. Гликолиз протекает в гиалоплазме (цитозоле) клетки. Гликолиз условно можно разбить на два этапа. В первом этапе происходит затрата энергии, второй этап, наоборот, характеризуется накоплением энергии в форме молекул АТФ. Основной путь катаболизма глюкозы у аэробных организмов – это аэробный распад. При гликолизе из глюкозы извлекается незначительная часть запасенной в ней энергии (2 молекулы АТФ на 1 молекулу глюкозы). Вся остальная энергия может быть получена при полном окислении глюкозы до СО2 и Н2О. Полное окисление осуществляется в ЦТК. Исходным веществом для цикла является ацетил–КоА. Для того, чтобы полностью окислить глюкозу необходимо конечный продукт аэробного гликолиза – ПВК – превратить в это исходное вещество. В аэробных условиях пировиноградная кислота, образовавшаяся при расщеплении глюкозы, не восстанавливается до лактата, а окисляется до ацетил–КоА. Этот путь называется окислительным декарбоксилированием ПВК. Промежуточные продукты анаэробного и аэробного непрямого распада до стадии образования ПВК идентичны. Глюконеогенез – это метаболический путь синтеза глюкозы из неуглеводных компонентов (ПВК, молочная кислота, глицерол, ряд аминокислот; такие аминокислоты называют гликогенными). Иными словами, предшественниками глюкозы в глюконеогенезе может быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов ЦТК.
Гликоген — полисахарид, образованный остатками глюкозы, связанными α-1→4 связями (α-1→6 в местах разветвления); основной запасной углевод человека и животных. является основной формой хранения глюкозы в животных клетках. Откладывается в виде гранул в цитоплазме во многих типах клеток. Гликоген образует энергетический резерв, который может быть быстро мобилизован при необходимости восполнить внезапный недостаток глюкозы. Только гликоген, запасённый в клетках печени (гепатоциты) может быть переработан в глюкозу для питания всего организма, при этом гепатоциты способны накапливать до 8 процентов своего веса в виде гликогена, что является максимальной концентрацией среди всех видов клеток. Общая масса гликогена в печени может достигать 100—120 граммов у взрослых. В мышцах гликоген перерабатывается в глюкозу исключительно для локального потребления и накапливается в гораздо меньших концентрациях (не более 1 % от общей массы мышц), в то же время его общий мышечный запас может превышать запас, накопленный в гепатоцитах. Небольшое количество гликогена обнаружено в почках, и ещё меньшее — в определённых видах клеток мозга (глиальных) и белых кровяных клетках.
7 , 8
Гликогенез распад гликогена в тканях
Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени – и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос-фат. Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглю-козу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорила-за). На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)-связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Реакция, катализируемая гликогенсинтазой. Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ, и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала. Распад гликогена печени служит в основном для поддержания уровня глюкозы в крови в постабсорбтивном периоде. Поэтому содержание гликогена в печени изменяется в зависимости от ритма питания. При длительном голодании оно снижается почти до нуля. Гликоген мышц служит резервом глюкозы - источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови. Как уже упоминалось ранее, в клетках мышц нет фермента глюкозо-6-фосфатазы, и образование свободной глюкозы невозможно. Расход гликогена в мышцах зависит в основном от физической нагрузки
9,10
Гликогеноз- накопление гликогена в органах и тканях. Заболевания относят к наследственным, вызванным нарушением активности одного из ферментов, участвующих в обмене гликогена.три основные формы заболевания:1) печеночную; 2) мышечную; 3) генерализованную (смешанную).
Печеночные гликогенозы : гликогеноз I типа или болезнь фон Гирке; гликогеноз III типа или болезнь Форбса-Кори; гликогеноз IV типа болезнь Андерсена.
Мышечные гликогенозы :Гликогеноз V типа болезнь Мак-Ардля.
Смешанные гликогенозы :Гликогеноз II типаболезнь Помпе.
Гликогеноз I типа или болезнь фон Гирке - обусловлен аутосомно-рецессивным дефектом глюкозо-6-фосфатазы. характеризуется недостатком или отсутствием фермента глюкозо-6-фосфатазы в печени и почках, вследствие чего гликоген не расщепляется и накапливается в этих органах. Организм удовлетворяет свои энергетические потребности путем усиления обмена жиров, что ведет к гиперлипидемии, жировой дегенерации печени, почек, ксантоматозу. Заболевание проявляется сразу после рождения или в грудном возрасте отсутствием аппетита, рвотой, понижением веса, гипогликемическими судорогами, комой. Печень увеличена, плотная на ощупь.Болезнь Мак Ардля обусловлена отсутствием миофосфорилазы. Активность печеночной фосфорилазы не изменена. В печени не происходит отложения гликогена. Поэтому нагрузка адреналином и глюкагоном дает нормальную реакцию. Характеризуется гипотонией мышц и их спазмами, наступающими при физических усилиях. Быстро возникает утомление больных. генерализованный, Помпе болезнь обусловленный дефицитом -глюкозидазы; характеризуется развитием сердечной недостаточности, адинамии, мышечной гипотонии, задержкой роста, расстройствами функций ц. н. с.; наследуется по аутосомно-рецессивному типу. Агликогеноз— наследственное заболевание, связанное с отсутствием фермента, ответственного за синтез гликогена — гликогенсинтетазы. При агликогенозе в печени полностью отсутствует гликоген, наблюдается полный дефицит гликогенсинтетазы при нормальной активности других ферментов, участвующих в обмене гликогена
Сахар крови норма 3,3–5,5 ммоль/л – норма независимо от возраста; сахарная кривая, отражающая изменения концентрации глюкозы в крови после сахарной нагрузки. Это довольно старое понятие. Сейчас этот анализ называется глюкозотолерантный тест. Глюкозотолерантный тест проводится пациентам, если концентрация сахара в крови натощак составляет от 5,7 до 6,9 ммоль/л, а также, если имеются факторы риска развития сахарного диабета (избыточный вес, гипертоническая болезнь, рождение крупного плода и наличие сахарного диабета у близких родственников). Построение сахарной кривой.В начале определяют содержание сахара в крови натощак. Затем исследуемому больному дают выпить раствор 50 г глюкозы в 200 мл воды. В течение последующих 3 ч берут пробы крови каждые 30 минут У здорового человека содержание сахара в крови после такой нагрузки увеличивается в течение первого часа примерно на 50% от исходного уровня, но не более 9,4 ммоль/л (170мг %), а ко второму часу снижается до начальной величины или даже ниже ее вследствие реактивного выброса инсулина. По показателям содержания глюкозы в крови и вычерчивается сахарная кривая. Диагностическая ценность теста заключается в том, что с его помощью можно диагностировать не только сахарный диабет, но и нарушение толерантности к глюкозе (латентный диабет), при котором сахар крови не повышается натощак, нет глюкозы в моче и отсутствуют клинические проявления. В таком случае, уровень сахара измеряется дважды: натощак, и после приема 75 грамм глюкозы (1,75 мг/кг для детей).
12 Вопр нейро-гуморальные механизмы поддержания уровня сахара крови
При избытке сахара в крови нервная система стимулирует функцию внутрисекреторной части поджелудочной железы. Теперь в кровь поступает больше гормона инсулина, и лишний сахар под его влиянием откладывается в печени и мышцах в виде гликогена. При усиленной мышечной работе, когда повышается потребление сахара и в крови его становится недостаточно, усиливается деятельность надпочечников. Гормон надпочечников адреналин способствует превращению гликогена в сахар. Так нервная система, воздействуя на железы внутренней секреции, стимулирует или тормозит отделение ими биологически активных веществ. Влияния нервной системы осуществляются через секреторные нервы. Кроме того, нервы подходят к кровеносным сосудам эндокринных желез. Меняя просвет сосудов, они влияют на деятельность этих желез. Инсулин – единственный гормон гипогликемического действия (снижает уровень глюкозы). Адреналин, клюкагон, АКТГ, СТГ, глюкокортикоиды – гипергликемические гормоны(повышают уровень глюкозы). Почечный порог Уровень вещества в крови, например сахара (глюкозы), превышение которого приводит к появлению этого вещества в моче. Например, если уровень сахара крови не превышает 160-170 мг%, то он не появляется в моче. Если сахар обнаруживается в моче, то это указывает на то, что уровень сахара крови превысил 160-170 мг%. Этот уровень сахара крови (160-170 мг%) и является почечным порогом для сахара крови.
глюкозы окисление: идет 2 путями – 2/3 глюкозы окисляется гликолитическим путем. 1/3 глюкозы окисляется пентозофосфатным путем.Гликолитический путь окисления глюкозы осуществляется в анаэробных (собственно гликолиз) и аэробных условиях. Окисление в анаэробных условиях называется гликолизом. При гликолизе из 1 молекулы глюкозы образуется 2 молекулы лактата и 2 АТФ. При аэробном гликолизе 1 молекулы глюкозы образуется 36 или 38 АТФ, выделяется 6 СО2, 6 Н2О. Пентозофосфатный цикл осуществляется в анаэробных условиях, в результате этого процесса выделяется СО2 и НАДФН2, НАДФН2 являются источниками энергии для восстановительного синтеза
Анаэробный распад глюкозы происходит при недостаточном содержании кислорода, в клетках мышечной ткани животного организма. Данный путь распада называется дихотомическим, т.к. в процессе происходит образование двух молекул триоз, содержащих по 3 С-атома из одной молекулы гексозы (6 С-атомов). Конечный продукт анаэробного превращения глюкозы – молочная кислота. Гликолиз протекает в гиалоплазме (цитозоле) клетки. Гликолиз условно можно разбить на два этапа. В первом этапе происходит затрата энергии, второй этап, наоборот, характеризуется накоплением энергии в форме молекул АТФ.
1 реакцией гликолиза является фосфорилирование глюкозы, т.е. перенос остатка фосфорной кислоты на глюкозу за счет энергии АТФ с образованием глюкозо-6-фосфата. Реакция катализируется ферментом гексокиназой. Киназами называются ферменты, катализирующие перенос остатка фосфорной кислоты от АТФ к акцептору. Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии и может считаться практически необратимым процессом.
2 реакцией гликолиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфат-изомеразы во фруктозо-6-фосфат (изомеризация альдозы в кетозу. Эта реакция протекает легко в обоих направлениях.
3 реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ. Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируется АТФ и стимулируется АМФ. Так, в неработающей мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно высокая. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза.
4 реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бифосфат расщепляется на две фосфотриозы. Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры peакция сдвигается в сторону большего образования триозофосфатов (дигидроксиацетонфосфата и глицеральдегид-3-фосфата (фосфоглицериновый альдегид)).
5 реакция - это реакция изомеризации триозофосфатов. Kaтализируется ферментом триозофосфатизомеразой.Дальнейшим превращениям будет подвергаться только глицеральдегид-3-фосфат.
В результате 6 реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается окислению с образованием 1,3- бифосфоглицериновой кислоты и восстановленной формы НАДН. С данной реакции начинается второй этап гликолиза.
7 реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфоглицериноой кислоты (3-фосфоглицерат). Это первая реакция гликолиза, в которой происходит образование АТФ (пример субстратного фосфорилирования).
8 реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат).
9 реакция катализируется ферментом енолазой, при этом фосфоглицериновая кислота в результате отщепления молекулы переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической.
10 реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстатное фосфолирование). Катализируется ферментом пируваткиназой.
В результате 11 реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН, образовавшегося в шестой реакции.
Конечным акцептором электронов в ходе гликолиза является ПВК, которая восстанавливается в молочную кислоту (лактат). Поэтому для нормального протекания этого метаболического пути требуется только глюкоза, которая может быть запасена клеткой в виде резервных полисахаридов в избыточном количестве.

Энергетический баланс гликолиза — две молекулы АТФ на одну молекулу глюкозы. На I этапе гликолиза расходуются две молекулы АТФ для активирования субстрата (в гексокиназной и фосфофруктокиназной реакциях). На II этапе образуются четыре молекулы АТФ (в фосфоглицераткиназной и пируваткиназной реакциях). Синтез АТФ осуществляется путем субстратного фосфорилирования.
Ключевые ферменты гликолиза:
1. Гексокиназа — это регуляторный фермент гликолиза во внепеченочных клетках. Гексокиназа аллостерически ингибируется глюкозо-6-фосфатом. Глюкокиназа — регуляторный фермент гликолиза в гепатоцитах. Синтез глюкокиназы индуцируется инсулином.
2. Фосфофруктокиназа-1. Это главный ключевой фермент, катализирует реакцию, лимитирующую скорость всего процесса (наиболее медленная реакция). Синтез фермента индуцируется инсулином. Аллостерические активаторы — АМФ, АДФ, фруктозо-2,6- дифосфат. Уровень фруктозо-2,6-дифосфата увеличивается под действием инсулина и понижается под действием глюкагона. Аллостерические ингибиторы — АТФ, цитрат.
3. Пируваткиназа. Фермент активен в нефосфорилированной форме. Глюкагон (в гепатоцитах) и адреналин (в миоцитах) стимулируют фосфорилирование фермента, а значит инактивируют фермент. Инсулин, наоборот, стимулирует дефосфорилирование фермента, а значит активирует фермент. Аллостерический активатор — Фр-1,6-ФФ. Аллостерический ингибитор — АТФ, ацетил КоА. Синтез фермента индуцирует инсулин.
Биологическая роль гликолиза:
1. Генерирование АТФ. Гликолиз — единственный процесс в клетках, продуцирующий АТФ без потребления кислорода. Клетки, имеющие мало или не имеющие вообще митохондрий, получают АТФ только в ходе гликолиза.
Значение гликолиза для эритроцитов. Гликолиз — единственный процесс, продуцирующий АТФ в эритроцитах и поддерживающий их целостность и функции.
Наследственный дефект пируваткиназы сопровождается гемолитической анемией. При этой патологии эритроциты имеют от 5 до 25 % нормальной пируваткиназной активности и, следовательно, скорость гликолиза низкая.
Промежуточный продукт гликолиза в эритроцитах — 2,3-дифосфоглицерат (2,3-ДФГ)
— понижает сродство гемоглобина к кислороду, способствуя диссоциации кислорода из оксигемоглобина и переходу его в ткани. Нарушения гликолиза в эритроцитах могут оказывать влияние на транспорт кислорода. Так, при недостаточности гексокиназы наблюдается понижение уровня 2,3-ДФГ и ненормально высокое сродство гемоглобина к кислороду. И наоборот, при недостаточности пируваткиназы содержание 2,3-ДФГ вдвое превышает норму, что обусловливает низкое сродство гемоглобина к кислороду.
2. Является источником углеводородных радикалов для процессов биосинтеза в клетках
16.
Гликогенолиз-биохимическая реакция, протекающая главным образом в печени и мышцах, во время которой гликоген расщепляется до глюкозы и глюкозо-6-фосфата. Гликогенолиз стимулируется гормонами глюкагоном и адреналином.
Расщепление гликогена включает несколько этапов. Сначала фосфорилаза последовательно отщепляет остатки глюкозы от концов боковых цепей гликогена При этом фосфорилируются альфа-1,4-связи и образуются молекулы глюкозо-1-фосфата . Фосфорилаза атакует боковую цепь до тех пор, пока не дойдет до точки, отстоящей на 4 остатка глюкозы от места ветвления (т. е. от альфа-1,6-связи). Затем вступает в действие система отщепления боковых цепей гликогена. Первый фермент этой системы - 4-альфа-D-глюканотрансфераза - отщепляет 3 из 4 остатков глюкозы и переносит их на свободный конец другой боковой цепи. Второй фермент - амило-1,6-глюкозидаза - отщепляет от главной цепи четвертый остаток глюкозы. После этого главная цепь гликогена становится доступной для фосфорилазы. В реакции, катализируемой амило-1,6-глюкозидазой, образуется глюкоза .
У здоровых людей при голодании до 8% гликогена печени расщепляется амило-1,6-глюкозидазой до глюкозы, а 92% гликогена расщепляется фосфорилазой до глюкозо-1-фосфата. Под действием фосфоглюкомутазы глюкозо-1-фосфат превращается в глюкозо-6-фосфат, при гидролизе которого в печени образуется глюкоза , поступающая в кровь.
Таким образом, основное количество глюкозы при голодании образуется в печени из глюкозо-6-фосфата.
17.
Спиртовое брожение — химическая реакция брожения, осуществляемая дрожжами, в результате которой одна молекула глюкозы преобразуется в 2 молекулы этанола и в 2 молекулы углекислого газа.
Реакция спиртового брожения подобна гликолизу. Расхождение начинается только после образования пирувата.Конечный этап гликолиза заменяется двумя ферментативными реакциями. Сначала пируват подвергается декарбоксилированию, продуктом которого является ацетальдегид. Данная реакция происходит при участии пируватдекарбоксилазы, ТПФ и ионов магния.

После ацетальдегид восстанавливается водородом, который отщепляется от кофермента НАДН. При этом ацетальдегид восстанавливается до этанола. Собственно, цель спиртового брожения — это окисление NADH, чтобы он мог снова принять участие в гликолизе. Катализатором является алкогольдегидрогеназа.
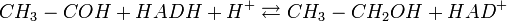
Таким образом, продуктами спиртового брожения являются этанол и , а не молочная кислота, как в молочнокислом брожении.
В результате получается реакция:

Применение спиртового брожения сходно с применением дрожжей: пивоварение, квасоварение, приготовление дрожжевого теста, виноделие и производство других алкогольных напитков.
18.
Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающий в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки.
1. Этапы аэробного гликолиза
В аэробном гликолизе можно выделить 2 этапа.
Подготовительный этап, в ходе которого глюкоза фосфорилируется и расщепляется на две молекулы фосфотриоз. Эта серия реакций протекает с использованием 2 молекул АТФ.
Этап, сопряжённый с синтезом АТФ. В результате этой серии реакций фосфотриозы превращаются в пируват. Энергия, высвобождающаяся на этом этапе, используется для синтеза 10 моль АТФ.
2. Реакции аэробного гликолиза
Превращение глюкозо-6-фосфата в 2 молекулы глицеральдегид-3-фосфата
Глюкозо-6-фосфат, образованный в результате фосфорилирования глюкозы с участием АТФ, в ходе следующей реакции превращается в фруктозо-6-фосфат. Эта обратимая реакция изомеризации протекает под действием фермента глюкозофосфатизомеразы.
Затем следует ещё одна реакция фосфорилирования с использованием фосфатного остатка и энергии АТФ. В ходе этой реакции, катализируемой фосфофруктокиназой, фруктозо-6-фосфат превращается в фруктозо-1,6-бисфосфат. Данная реакция, так же, как гексокиназная, практически необратима, и, кроме того, она наиболее медленная из всех реакций гликолиза. Реакция, катализируемая фосфофруктокиназой, определяет скорость всего гликолиза, поэтому, регулируя активность фосфофруктокиназы, можно изменять скорость катаболизма глюкозы.
Фруктозо-1,6-бисфосфат далее расщепляется на 2 триозофосфата: глицеральдегид-3-фосфат и дигидроксиацетонфосфат. Реакцию катализирует фермент фруктозобисфосфатальдолаза, или просто альдолаза. Этот фермент катализирует как реакцию альдольного расщепления, так и альдольной конденсации, т.е. обратимую реакцию. Продукты реакции альдольного расщепления - изомеры. В последующих реакциях гликолиза используется только глицеральдегид-3-фосфат, поэтому дигидроксиацетонфосфат превращается с участием фермента триозофосфатизомеразы в глицероальдегид-3-фосфат.
В описанной серии реакций дважды происходит фосфорилирование с использованием АТФ. Однако расходование двух молекул АТФ (на одну молекулу глюкозы) далее будет компенсировано синтезом большего количества АТФ.
Пути катаболизма глюкозы:


Превращение глюкозо-6-фосфата в триозофосфаты.
19. Окисление цитоплазматического NADH
в митохондриалъной дыхательной цепи.
Челночные системы
NADH, образующийся при окислении глицеральдегид-3-фосфата в аэробном гликолизе, подвергается окислению путём переноса атомов водорода в митохондриальную дыхательную цепь. Однако цитозольный NADH не способен передавать водород на дыхательную цепь, потому что
электроны с FADH2 переходят на убихинон и далее по ЦПЭ
Глицеролфосфатная челночная система работает в клетках белых мышц и гепатоцитов. Однако в клетках сердечных мышц митохондриальная глицерол-3-фосфатдегидрогеназа отсутствует. Вторая челночная система, в которой участвуют малат, цитозольная и митоховдриальная малат-дегидрогеназы, является более универсальной. В цитоплазме NADH восстанавливает оксалоа-цетат в малат, который при участии переносчика проходит в митохондрии, где окисляется в оксалоацетат NAD-зависимой маЛатдегидрогеназой. Восстановленный в ходе этой реакции NAD отдаёт водород в митоховдриальную ЦПЭ. Однако образованный из малата оксалоацетат выйти самостоятельно из митохондрий в цитозоль не может, так как мембрана митохондрий для него непроницаема. Поэтому оксалоацетат превращается в аспартат, который и транспортируется в цитозоль, где снова превращается в оксалоацетат. Превращения оксалоацетата в аспартат и обратно связаны с присоединением и отщеплением аминогруппы (реакции трансаминирования. Эта челночная система называется малат-аспартатной. Результат её работы - регенерация цитоплазматического NAD+ из NADH.
Обе челночные системы существенно отличаются по количеству синтезированного АТФ. В первой системе соотношение Р/О равно 2, так как водород вводится в ЦПЭ на уровне KoQ. Вторая система энергетически более эффективна, так как передаёт водород в ЦПЭ через митохондриальный NAD+ и соотношение Р/О близко к 3. Митоховдриальная мембрана для него непроницаема. Перенос водорода через мембрану происходит с помощью специальных систем, называемых "челночными". В этих системах водород транспортируется через мембрану при участии пар субстратов, связанных соответствующими дегидрогеназами, т.е. с обеих сторон митохондри-альной мембраны находится специфическая дегидрогеназа. Известны 2 челночные системы. В первой из этих систем водород от NADH в цитозоле передаётся на дигидроксиацетонфосфат ферментом глицерол-3-фосфатдегидрогеназой (NAD-зависимый фермент, назван по обратной реакции). Образованный в ходе этой реакции глицерол-3-фосфат, окисляется далее ферментом внутренней мембраны митохондрий - глицерол-3-фосфатдегидрогеназой (FAD-зависимым ферментом).
20. Энергетическая ценность аэробного дихотомического окисления глюкозы.
Выход АТФ при аэробном гликолизе
На образование фруктозо-1,6-бисфосфата из одной молекулы глюкозы требуется 2 молекулы АТФ (реакции 1 и 3 на рис. 7-33). Реакции, связанные с синтезом АТФ, происходят после распада глюкозы на 2 молекулы фосфотриозы, т.е. на втором этапе гликолиза. На этом этапе происходят 2 реакции субстратного фосфорилирования и синтезируются 2 молекулы АТФ (реакции 7 и 10). Кроме того, одна молекула глицеральдегид-3-фосфата дегидрируется (реакция 6), a NADH передаёт водород в митохондриальную ЦПЭ, где синтезируется 3 молекулы АТФ путём окислительного фосфорилирования. В данном случае количество АТФ (3 или 2) зависит от типа челночной системы. Следовательно, окисление до пирувата одной молекулы глицеральдегид-3-фосфата сопряжено с синтезом 5 молекул АТФ. Учитывая, что из глюкозы образуются 2 молекулы фосфотриозы, полученную величину нужно умножить на 2 и затем вычесть 2 молекулы АТФ, затраченные на первом этапе. Таким образом, выход АТФ при аэробном гликолизе составляет (5×2) - 2 = 8 АТФ.
Эффект Пастера.
Это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода. Биохимический механизм эффекта заключается в конкуренции за пируват между пируватдегидрогеназой, превращающей пируват в ацетил-S-КоА, и лактатдегидрогеназой, превращающей пируват в лактат.
У пируватдегидрогеназы сродство гораздо выше и в обычных аэробных условиях она окисляет большую часть пировиноградной кислоты. Как только поступление кислорода уменьшается (недостаток кровообращения, тромбоз и т.п.) происходит следующее:
внутримитохондриальные процессы дыхания не идут и НАДН в дыхательной цепи не окисляется,
моментально накапливающийся в митохондриях НАДН тормозит цикл трикарбоновых кислот,
ацетил-S-КоА не входит в ЦТК и ингибирует ПВК-дегидрогеназу.
В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную.
При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.
21.
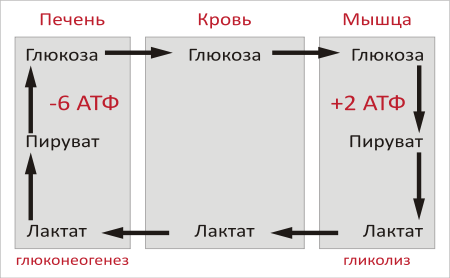
Цикл Кори— совокупность биохимических ферментативных процессов транспорта лактата из мышц в печень, и дальнейшего синтеза глюкозы из лактата, катализируемое ферментами глюконеогенеза.
Биологический смысл
При интенсивной мышечной работе, а также в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах или мышцах) глюкоза вступает на путь анаэробного гликолиза с образованием лактата. Лактат не может далее окисляться, он накапливается (при его накоплении в мышцах раздражаются чувствительные нервные окончания, что вызывает характерное жжение в мышцах). С током крови лактат поступает в печень. Печень является основным местом скопления ферментов глюконеогенеза (синтез глюкозы из неуглеводных соеднений), и лактат идет на синтез глюкозы.
Реакция превращения лактата в пируват катализируется лактатдегидрогеназой, далее пируват подвергается окислительному декарбоксилированию или может подвергаться брожению.
22.
ГЛЮКОНЕОГЕНЕЗ- синтез моносахаридов (гл. обр. глюкозы) из неуглеводных предшественников, происходящий в живых клетках под действием ферментов.
|
из
5.00
|
Обсуждение в статье: Превращение глюкозо-6-фосфата в триозофосфаты |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы