 |
Главная |


Andio R., D. Silvera, S. D. Suggett, P. L. Achacoso, 9 страница
|
из
5.00
|
Сидерофоры
Железо — один из наиболее распространенных на Земле элементов, абсолютно необходимых живым организмам. Однако в той форме, в какой железо присутствует в почве, оно не может прямо использоваться микроорганизмами. Дело в том, что его преобладающей природной формой являются трехвалентные ионы. Их растворимость очень мала— при pH 7,4 она равна примерно 10~18 М, и этого количества абсолютно недостаточно для поддержания роста микроорганизмов. Чтобы выжить в таких условиях, почвенные микроорганизмы синтезируют и секретируют небольшие низкомолекулярные железосвязывающие соединения мол. массой примерно 400—1000 Да, известные под названием сидеро-форов (рис. 14.8). Они эффективно связывают Fe(IlI) и транспортируют его к клеткам микроорганизмов, где оно связывается с клеточными рецепторами и попадает внутрь клеток. Здесь железо высвобождается и может использоваться микроорган измом.
Бактерии, стимулирующие рост растений, подавляют пролиферацию фитопатогенных грибов, синтезируя сидерофоры, которые связывают большую часть Fe(lII), находящегося в слое почвы, непосредственно прилегающем к корню растения (в ризосфере). Фитопатогенные грибы тоже синтезируют сидерофоры, но они обычно обладают более низким сродством к железу, чем сидерофоры, синтезируемые стимулирующими рост растений бактериями. Это позволяет последним одерживать верх в конкурентной борьбе с фито патогенны ми грибами за имеющееся железо.
В отличие от фитопатогенных микроорганизмов, растения, как правило, не страдают от локального истощения железа в почве в результате поглощения его бактериями, стимулирующими рост растений. Большинство растений могут расти при значительно меньших концентрациях железа, чем микроорганизмы. Кроме того, есть данные, что железо, связанное бактериальными сидерофорами, может ассимилироваться растениями и использоваться ими для своих нужд.
Поскольку связывание железа бактериальными сидерофорами может одновременно приводить к подавлению пролиферации самых
322 ГЛАВА 14

|
| Рис. 14.8. Структура сидерофора псевдобактина, продуцируемого штаммом Pseudomonas Β10. С одной молекулой сидерофора связан один ион Fe(III). |
разных фитопатогенных микроорганизмов, исследуется возможность использования их для создания более эффективных систем биоконтроля.
Многие стимулирующие рост растений флуоресцирующие псевдомонады секретируют сидерофор, представляющий собой линейный гексапептид, который состоит из чередующихся L-и D-аминокислот и связанного флуоресцентного хромофора (рис. 14.8). Один из таких сидерофоров, так называемый псевдобактин, обладает сродством к Fe(III) ≅ 1025 л · моль~1. Сходные сидерофоры синтезируют все флуоресцирующие псевдомонады.
Предприняты также первые попытки исследовать синтез псевдобактина у стимулирующей рост растений бактерии Pseudomonas putida WCS358. С помощью мутагенеза были получены 28 мутантов этого микроорганизма, не способных синтезировать сидерофор. Их отбор осуществляли по: 1) отсутствию флуоресценции в УФ-свете; 2) неспособности к росту в присутствии дипиридила — вещества, связывающего большую часть железа в культуральной среде. При очень малых концентрациях ионов железа растут только те клетки, которые синтезируют сидерофоры. Был создан банк клонов ДНК P. putida WCS358 с помощью космидного вектора pLAFRl с широким кругом хозяев и путем конъюгации осуществлена трансформация всех 28 мутантных форм. Трансформанты были проверены на способность к флуоресценции в УФ-свете и/или способность к росту в присутствии дипиридила. Идентифицированы тринадцать
разных комплементирующих космидных клонов со средним размером вставки 26 т. п. н. Детальные исследования показали, что эти клоны соответствуют по крайней мере пяти отдельным кластерам генов.
Один из этих кластеров был исследован более детально. Его минимальная длина составила примерно 33,5 т. п. н., он содержал пять оперо-нов по крайней мере с семью отдельными генами. Таким образом, как и фиксация азота и образование клубеньков, биосинтез сидерофоров -это сложный процесс. Поскольку каждый сидерофор кодируется несколькими генами, получение рекомбинантных бактерий, способных синтезировать модифицированный сидерофор, -задача не из легких. К счастью, есть другие способы повысить эффективность использования бактерий, стимулирующих рост растений, в качестве инструмента биоконтроля. Например, можно расширить круг узнаваемых одним бактериальным штаммом комплексов железо—сидерофор, так чтобы один рекомбинантный штамм мог узнавать и использовать сидерофоры, синтезируемые другими почвенными микроорганизмами, повышая тем самым свою конкурентоспособность. Для этого были клонированы гены рецепторов комплекса железо—сидерофор одной из бактерий, стимулирующих рост растений, и введены в другие штаммы.
Антибиотики
Один из наиболее эффективных механизмов, которые используют стимулирующие рост растений бактерии для подавления пролиферации
Бактерии, стимулирующие рост растений 323
фитопатогенов, — синтез антибиотиков. Так, псевдомонады, использующиеся для биоконтроля, синтезируют такие антибиотики, как агроцин 84, агроцин 434, 2,4-диацетилфлороглюцинол, гербиколин, оомицин, феназины, пиолутеорин и пирролнитрин.
Способность бактерий, стимулирующих рост растений, подавлять пролиферацию фитопатогенов можно повысить, если ввести в эти бактерии гены, кодирующие биосинтез антибиотиков, которые обычно синтезируются другими бактериями. Это позволит расширить спектр фитопатогенов, рост которых способна подавлять одна бактерия. Более того, ограничивая размножение других почвенных микроорганизмов, секретирующие антибиотик бактерии, стимулирующие рост растений, облегчают свою собственную пролиферацию, поскольку уменьшается число конкурентов за ограниченные пищевые ресурсы, а с помощью методов генной инженерии со временем удастся увеличить выход бактериальных антибиотиков.
Синтез большинства противогрибковых метаболитов, продуцируемых псевдомонадами, по-видимому, контролируется белком, который действует как общий регулятор транскрипции; следовательно, уровень синтеза антибиотика можно повысить изменением общей регуляции. Например, в случае Pseudomonas fluorescens СHАО его удалось повысить с помощью трансформации микроорганизма вектором, несущим ген «домашнего хозяйства» rpoD, который кодирует σ70-субъединицу РНК-полимеразы. Рекомбинантный штамм лучше защищал корни растений огурца от повреждений, вызываемых грибом Pythium ultimum (табл. 14.7). В другом исследовании синтез антибиотика пиолутеорина тем же микроорганизмом стимулировали инактивацией генов pqq, участвующих в биосинтезе пирролхинолинхинона — кофактора различных гидрогеназ. Механизм такой стимуляции до конца не установлен; возможно, в результате мутации поток метаболитов от других реакций биосинтеза переключается на биосинтез пиолутеорина.
В настоящее время на рынке имеется только один рекомбинантный микроорганизм, использующийся для биоконтроля: Agrobacterium radiobacter К84, Этот штамм продается в Австра-
| Таблица 14.7. Влияние дополнительных копий rpoD- гена Pseudomonas fluorescens CHA0 на способность этой бактерии защищать корни растений огурца от повреждений, вызываемых патогенным грибом Pythium ultiтит1) | |

| |
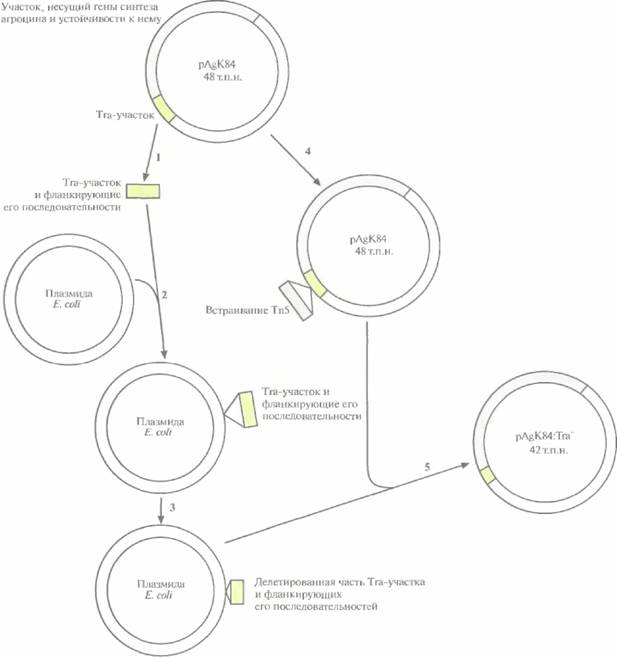
лии начиная с 1989 г. как средство борьбы с корончатым галлом — болезнью, которую вызывает Agrobacterium tumefaciens. Заболеванию подвержены миндаль и косточковые плодовые деревья, такие как персики. A. radiobacter синтезирует антибиотик агроцин 84, токсичный для A. tumefaciens. Однако при случайном попадании в него плазмиды от A. radiobacter, содержащей гены биосинтеза агроцина 84, могут возникнуть агроцинустойчивые штаммы Л. tumefaciens. Чтобы избежать этого, из плазмиды pAgK84, несущей гены биосинтеза агроцина 84, был удален участок, ответственный за перенос плазмиды (рис. 14.9). В результате штамм A. radiobacter больше не мог передавать рекомбинантную агроциновую плазмиду патогенным агробактериям, но сохранял способность к биоконтролю.
Ферменты
Некоторые бактерии, стимулирующие рост растений, синтезируют такие ферменты, как хитиназа, р-1,3-глюканаза, протеаза и липаза, которые разрушают клеточную стенку грибов. В одном из экспериментов удалось снизить частоту возникновения болезней, вызываемых фитопатогенными грибами Rhizoctonia solani, Sclerotium rolfsii и Pythium ultimum, с помощью штамма Pseudomonas cepacia, синтезирующего
|
| Рис. 14.9. Конструирование неспособной к переносу (Тrа~) модификации плазмиды pAgK84 A. radiobacter, кодирующей гены биосинтеза агроцина 84 и гены устойчивости к нему. Используя рестрикционную карту pAgK84, из плазмиды вырезали (1) фрагмент, который содержал участок Тга, ответственный за перенос, вместе с фланкирующими его последовательностями, и встроили его в плазмиду E. coli(2). С помощью рестриктазной обработки из клонированного фрагмента удалили примерно 80% Tra-участка и часть фланкирующих его последовательностей (в сумме примерно 6 т.п.н.) (3). Осуществили гомологичную рекомбинацию между плазмидой E. coli, содержащей «урезанный» Tra-участок, и плазмидой pAgK84, которая содержала встроенный в Tra-участок транспозон Тn5 с геном устойчивости к канамицину (4). Получили несколько производных плазмиды pAgK84 с частично удаленным Тга-участком (5), Тга~-плазмида pAgK84 больше не могла передаваться другой агробактерии в ходе конъюгации, но все еще была способна детерминировать синтез агроцина 84 и обеспечивать устойчивость к нему. Рисунок выполнен без соблюдения масштаба. |
Бактерии, стимулирующие рост растений 325
фермент β-1,3-глюканазу, который разрушал грибной мицелий. В ходе других исследований было показано, что противогрибковая активность трех штаммов Enterobacter agglomerans, стимулирующих рост растений, обусловлена наличием у них комплекса из четырех разных полипептидов, которые, действуя совместно, расщепляют хитин клеточной стенки грибов. Эти бактерии хорошо защищали растения хлопчатника от заражения Rhizoctonia solani. В то же время Тn5-мутанты Е. agglomerans, не продуцирующие активной хитиназы, не были способны защитить растения от патогенных грибов.
Многие бактериальные ферменты, разрушающие клеточную стенку грибов, в том числе хи-тиназа и ß-глюканаза, кодируются одним геном. Было бы разумно выделить эти гены и ввести их бактериям, стимулирующим рост растений, с тем чтобы получить штаммы, синтезирующие, например, и антибиотики, и ферменты, разрушающие клеточную стенку грибов. Были проведены эксперименты, в которых ген хитиназы, выделенный из бактерии Serratia marcescens, был перенесен в клетки Trichoderma harzianum и R. meliloti. Оба трансформированных микроорганизма синтезировали хитиназу и обладали повышенной противогрибковой активностью. При введении гена хитиназы S. marcescens в штамм P. fluorescens. стимулирующий рост растений, был получен трансформант, стабильно секретирующий хитиназу и эффективно подавляющий размножение фитопатогенного гриба Rhizoctonia solani.
Образование кристаллов льда и антифризные белки
Некоторые патогенные поражающие листья бактерии типа Pseudnmonas syringae синтезируют при низких температурах специфические белки, служащие центрами образования кристаллов льда на поверхности листа при температурах ниже нуля. По мере своего роста кристаллы прокалывают растительные клетки и необратимо повреждают растение, а бактерии получают в свое распоряжение питательные вещества, высвободившиеся из разрушенных растительных клеток. Если белки — центры кристаллизации на поверхности листа — отсутствуют, то непродолжительные ночные заморозки могут и не принести вреда растению, поскольку образование кристаллов льда в цитоплазме растительной клетки обычно начинается при температуре на несколько градусов ниже точки замерзания (т. е. происходит ее переохлаждение). Чтобы предотвратить кристаллизацию на листьях таких культур, как земляника, можно еше до заморозков распылить над растениями мутантные бактерии P. syringae, не способные синтезировать белки — центры кристаллизации. Такие мутантные формы могут быть созданы с помощью технологнии рекомбинантных ДНК или обычного мутагенеза с последующим отбором, и они при достаточной концентрации вытеснят бактерии дикого типа.
Одним из важных условий эффективности биоконтроля патогенных микроорганизмов с помощью бактерий, стимулирующих рост растений, является способность этих бактерий к распространению в естественных условиях. В Канаде, скандинавских странах и на севере США они должны сохранять жизнеспособность в условиях долгих холодных зим, а весной размножаться при относительно низких температурах почвы (~5—10 °С). Поскольку микроорганизмы используют разные адаптивные стратегии выживания в неблагоприятных условиях, можно попытаться сконструировать с помощью генной инженерии рекомбинантные бактерии, оптимально приспособленные к низким температурам. Недавно было показано, что некоторые почвенные бактерии (а среди них встречаются и такие, которые стимулируют рост растений) могут размножаться при 5 °С и секретировать в окружающую среду антифризные белки при низких температурах. Такие белки регулируют образование кристаллов льда внутри бактериальной клетки. Хотя в их присутствии кристаллы все же формируются, они не достигают больших размеров и не разрушают клетки. Как только будут идентифицированы гены бактериальных антифризных белков, их можно будет перенести в клетки бактерий, стимулирующих рост растений, с тем чтобы получить трансформированные бактерии, устойчивые к низким температурам. Пока нет никаких данных о наличии связи между антифризной активностью бактерий и механизмом, обеспечивающим их выживание при низких температурах. Очень ин-
326 ГЛАВА 14
тересно проверить, является ли синтез антифризного белка частью адаптивной стратегии, используемой некоторыми бактериями для обеспечения устойчивости к холоду.
Стимуляция роста растений свободноживущими бактериями
Бактерии, стимулирующие рост растений, оказывают свое действие несколькими способами: 1) фиксируют атмосферный азот, который затем используется растением; 2) синтезируют сидерофоры, которые солюбилизируют и связывают железо из почвы и обеспечивают им растительные клетки; 3) синтезируют фитогормоны, ускоряющие разные стадии роста; 4) солюбилизируют минеральные вещества (такие, как фосфор), которые затем используются растением; 5) синтезируют ферменты, способные регулировать уровень растительных гормонов. Каждая бактерия, стимулирующая рост растений, может использовать один или несколько из этих механизмов.
Фиксация азота вносит совсем небольшой вклад в тот положительный эффект, который дают бактерии, стимулирующие рост растений. Не все такие бактерии являются диазотрофами, а многие из них усваивают лишь ограниченное количество азота.
Для поглощения железа из почвы некоторые растения используют бактериальные комплексы железо—сидерофор; без этого их рост в большинстве случаев был бы сильно замедлен. Однако, несмотря на то что бактериальные сидерофоры несомненно вносят вклад в питание растений и, следовательно, в их рост, этот эффект, как правило, не очень велик.
Как именно бактерии, стимулирующие рост растений, способствуют поглощению растением таких минеральных веществ, как фосфор, до конца не установлено. Высказывалось предположение, что у растений, обработанных стимулирующими их рост бактериями, лучше развивается корневая система, а потому они более эффективно поглощают из почвы нужные им вещества, т. е. влияние бактерий носит опосредованный характер. Однако эксперименты с Azospirillum показали, что этот организм увеличивает поглощение именно минеральных веществ, возможно, синтезируя и секретируя органические кислоты, которые растворяют и связывают некоторые из этих веществ.
Очень часто различные эффекты бактерий, стимулирующих рост растений, объясняют способностью их к синтезу фитогормонов. Большинство исследований в этой области относится к выяснению роли одного из классов фитогормонов — ауксинов. Наиболее распространенный и лучше всего охарактеризованный ауксин это индолил-3-уксусная кислота (ИУК). Она стимулирует как быстрые ответы (например, удлинение растительных клеток), так и длительные (ускорение деления и диффе-ренцировки). Растения тоже могут синтезировать ауксин. Часто это не позволяет определить, какой именно ауксин дает необходимый эффект: бактериальный или растительный. Тем не менее можно утверждать, что бактерии, стимулирующие рост растений, оказывают свое действие именно через изменение гормонального баланса в растениях.
Недавно обнаружилось, что многие бактерии, стимулирующие рост растений, синтезируют фермент, способный регулировать уровень растительного гормона этилена. Этот фермент, 1-аминоциклопропан-1-карбоксилат(АЦК)—дезаминаза, гидролизует АЦК, который является непосредственным предшественником этилена при биосинтезе в растениях. Одно из объяснений роли этого фермента состоит в следующем. Бактерия связывается с оболочкой семени или с корнями растения, а затем поглощает и гидролизует АЦК, понижая концентрацию этилена в тканях растения. Во многих растениях этилен стимулирует прорастание семян и выводит их из состояния покоя; однако, если после прорастания уровень этилена оказывается слишком высоким, удлинение корней замедляется. Таким образом, бактериальная АЦК—дезаминаза предотвращает уменьшение-скорости роста корней, и растение развивается быстрее. Кроме того, многие бактерии, стимулирующие рост растений, синтезируют ИУК, а избыток ИУК, не израсходованный на стимуляцию удлинения растительных клеток или ускорение деления, активирует АЦК-синтазу, что приводит к повышению концентрации этилена. Присутствие активной АЦК—дезаминазы препятствует накоп-
Бактерии, стимулирующие рост растений 327

| Рис. 14. 10. Схематическое изображение механизма, с помощью которого бактерии, стимулирующие рост растений, снижают концентрацию этилена в растительных тканях и тем самым предотвращают ингибирование роста корней. Бактериальная клетка, прикрепившаяся к поверхности семени или корня развивающегося растения, синтезирует и секретирует индолил-3-уксусную кислоту (ИУК), стимулирующую рост растения. Попадая в растение, бактериальная ИУК (вместе с ИУК, синтезируемой самим растением) стимулирует либо деление растительной клетки и ее удлинение, либо фермент АЦК-синтазу, который катализирует превращение S-аденозилметионина (AdoMet) в АЦК. Значительная часть АЦК вместе с другими малыми молекулами, обычно содержащимися в семенном или корневом эксудате, экскретируется корнями растения или семенами, поглощается бактерией и гидролизуется АЦК-дезаминазой до аммиака и α-кетобутирата. В результате количество АЦК вне растения снижается. Чтобы сохранить равновесие между АЦК внутри и снаружи, растение секретирует его больше. Соответственно его концентрация, а следовательно, и концентрация этилена в растительных тканях уменьшается. (Из работы Glick et al., J. Theor. Вiol., in press.) |
лению АЦК даже при высоких концентрациях ИУК,так что концентрация этилена не повышается до уровня, при котором замедляется рост растения (рис. 14.10). После детального изучения механизмов, с помощью которых бактерии, стимулирующие рост растении, оказывают свое действие, появится возможность создавать рекомбинатные микроорганизмы, способные стимулировать рост самых разных растений в различных условиях.
ЗАКЛЮЧЕНИЕ
Многие почвенные микроорганизмы обладают способностью стимулировать рост растений. Были исследованы молекулярные механизмы, лежащие в основе этой стимуляции, с тем чтобы выяснить, можно ли использовать полезные почвенные бактерии вместо химических удобрений. Полезные бактерии могут оказывать свое влияние непосредственно, поставляя растениям фиксированный азот, хелатированное железо, фитогормоны или облегчая поглощение ими фосфора. Но влияние может быть и опосредованным, через подавление роста фитопатогенных микроорганизмов.
Из всех бактерий, стимулирующих рост растений и уже использующихся в сельском хозяйстве, наиболее детально изучены члены семейства Rhizobium и Bradyrhizobium. Эти микроорганизмы вступают в сложные облигатные симбиотические отношения со строго определенными растениями.
Молекулярные основы фиксации азота всесторонне исследовались на К. pneumoniae, которая может служить модельной системой для изучения симбиотических бактерий семейств Rhizobium и Bradyrhizobium. Детально охарактеризована нитрогеназа, азотфиксирующий фермент. Молекулярно-генетические исследования показали, что фиксация азота бактериями — это сложный процесс; в нем участвует семь координирование регулируемых оперонов, кодирующих в общей сложности 20 разных белков. Это делает пока невозможным создание с помощью методов генной инженерии растений, которые могли бы сами усваивать азот, и других азотфиксирующих бактерий.
Азотфиксирующий фермент нитрогеназа, используя энергию гидролиза АТР, катализирует образование газообразного водорода (Н2). Некоторые штаммы Rhizobium синтезируют фермент гидрогеназу. Он катализирует превращение in vivo H, в Н+, что увеличивает эффек-
328 ГЛАВА 14
тивность фиксации азота. Если штамм содержит неактивную гидрогеназу, его способность фиксировать азот и стимулировать рост растения уменьшается, С учетом всего сказанного выше были предприняты попытки ввести клонированные гены гидрогеназ в штаммы Rhizobium, вступающие в симбиотические отношения с сельскохозяйственными культурами. По предварительным данным, проводя генноинженерные модификации генов гидрогеназ, можно создать штаммы Rhizobium, обладающие более высокой способностью стимулировать рост растений.
Вступая в симбиотические отношения с растениями, штаммы Rhizobium стимулируют образование на их корнях клубеньков, где и происходит размножение этих бактерий и фиксация азота. Разумно было предположить, что, если с помощью методов генной инженерии удастся создать бактерии, способствующие образованию большего количества клубеньков, конкурентоспособность инокулирующих штаммов Rhizobium в борьбе за место на корнях растений-симбионтов повысится по сравнению со штаммами дикого типа, К сожалению, обнаружилось, что в образовании клубеньков участвует множество разных генов, и эта сложность затрудняет проведение соответствующих молекулярно-генетических экспериментов.
Опосредованная стимуляция роста растений бактериями состоит в защите растений от повреждений, вызываемых фитопатогенными грибами или бактериями. Такая зашита осуществляется при участии специфических соединений, синтезируемых бактериями, которые стимулируют рост растений: сидерофоров, антибиотиков, других малых молекул и различных ферментов. Некоторые другие продукты синтеза, в частности фитогормоны и АЦК—дезаминаза, влияют на рост растений непосредственно. Есть надежда, что когда-нибудь гены биосинтеза всех перечисленных соединений можно будет использовать для создания бактерий — более эффективных стимуляторов роста растений.
ЛИТЕРАТУРА
Adams M. W. W., L. E. Mortenson, J.S. Chen.1981. Hydrogenase. Biochitn, Biophys. Acta 594: 105-176.
Albrecht S. L.f R. J. Maier, F. J. Hanus, S. A. Russell, D. W. Emerich, H. J.Evans. 1979, Hydrogenase in Rhizobium japonicum increases nitrogen fixation by nodulated soybeans. Science 203: 1255-1257.
Arp D. J. 1990. H2 cycling in N2 fixation: past, present, and future outlook, p. 67—76. In P. M. Gresshoff, L E. Roth, G. Stacey, W. E. Newton (ed.), Nitrogen Fixation: Achievements and Objectives. Chapman & Hall, New York, N.Y.
Bar-Ness E.f Y. Chen, Y.Hadar, H.Marschner, V. Römheld.1991. Siderophores of Pseudotnonas puttda as an iron source for dicot and monocot plants, p. 271-281. In Y. Chen, Y. Hadar (ed.), Iron Nutrition ana Interactions in Plants. Kluwer Academic Publishers, Dordrecht, The Netherlands.
Cantrell M. A., R. A. Haugland, H. J. Evans.1983. Construction of a Rhizobium japonicum gene bank and use in isolation of a hydrogen uptake gene. Proc. Natl. Acad. Sei. USA 80: 181-185.
Chet I., J. Inbar.1994, Biological control of fungal pathogens, Appl Biochem. Biotechnol. 48: 37-43.
Crosa J. H.1989. Genetics and molecular biology of siderophore-mediated iron transport in bacteria. Microbioi. Rev. 53: 517-530.
Evans H. J., A. R. Harker, H. Papen, S. A. RusseU, F. J. Hanus, M. Zuber.1987. Physiology, biochemistry, and genetics of the uptake hydroge-nase in Rhizobia. Annu. Rev. Microbioi. 41: 335-361.
Fischer H.-M.1994. Genetic regulation of nitrogen fixation in Rhizobia. Microbioi. Rev. 58: 352-386.
Glick B. R1995. The enhancement of plant growth by free-living bacteria. Can. J. Kficrvbioi. 41:109-117.
Glick B. R., Y.Bashan. 1997. Genetic manipulation of plant growth-promoting bacteria to enhance biocontrol of phytopathogens. Biotechnol. Adv. 15:353-378.
Glick B. R., C. B. Jacobson, M. M. K.Schwar/c, J. J. Pasternak.1994. l-Aminocyclopropane-1-carboxylic acid deaminase mutants of the plant growth-promoting rhizobacterium Pseudomonas putida GR12-2 do not stimulate canola root elongation. Can. J. Microbioi. 40: 911-915.
Glick B. R., D. M.Penrose, J. Li. A model for the lowering of plant ethylene concentrations by
Бактерии, стимулирующие рост растений 329
plant growth-promoting bacteria. / Theor. ВЫ., in press.
Glick B. R.,.). Zeisler, A. M. Banaszuk, .1. D. Friesen, W. G. Martin. 1981. The identification and partial characterization of a plasmid containing the gene for the membrane-associated hydrogenase from E. colt. Gene 15: 201-206.
Gresshoff P. IVL, L. E. Roth, G. Stacey, W. E. Newton (ed.). 1990. Nitrogen Fixation: Achievements and Objectives. Chapman & Hall, New York, N.Y.
Hennecke H.1990. Nitrogen fixation genes involved in the Bradyrhizobium japonicum-soybean symbiosis. FEBS Lett. 268: 422-426.
Higashi S. 1993. (/^йф>)КА/доем/т-р1ап1 communications involved in infection and nodulation. J. Plant Res. 106:201-211.
Jones D. A., M, H. Ryder, B. G. Clare, S. K. Farrand, A, Kerr.1988. Construction of a Tra — deletion mutant of a pAgK84 to safeguard the biological control of crown gall. Mol. Gen. Genet. 212:207-214.
Kloepper J. W., R. Lifshitz, M. N. Schroth.1988. Pseuâomonas inoculants to benefit plant production. 1SI Atlas Sei. Anim. Plant Set. 60-64.
Layva A-, J. M. Palacios, T. Mozo, T.Ruiz-Argüeso.
1987. Cloning and characterization of hydrogen uptake genes from Rhizobium leguminosarum. J. Bacterial. 169: 4929-4934.
Lerouge P., P. Roche, С Faucher, F.Maillet, G. Truchet, J, C.Promé, J. Dénaric.1990. Symbiotic host-specificity of Rhizobium melüoti is determined by a su 1 ρ hated and асу la ted oligosaccharide signal. Nature 344: 781—784.
Long S. R., \V. J. Buikema, F. M.Ausübet. 1982. Cloning of Rhizobium meliloti nodulation genes by direct complementation of Nod~ mutants. Nature 298: 485-488.
Lynch J. M.1990. Beneficial interactions between microorganisms and roots. Biotechnol. Adv. 8: 335-346.
Marugg J. D., M. van Spanje, W. P. M. Hoekstra, B.Schippers, P. J. Weisbeek. 1985. Isolation and analysis of genes involved in siderophore biosynthesis in plant-growth-stimulating Pseudomonas putida WC358. /. BacterioL 164:563-570.
Marugg J. D., H. B. Nielander, A. J. G. Horrevoets, I. van Megen, I, van Genderen, P. J.Weisbeek.
1988. Genetic organization and transcriptional
analysis of a major gene cluster involved in siderophore biosynthesis in Pseudomonas putida WCS358. J. BacterioL 170: 1812-1819.
|
из
5.00
|
Обсуждение в статье: Andio R., D. Silvera, S. D. Suggett, P. L. Achacoso, 9 страница |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы