 |
Главная |


Голантарктическое царство
|
из
5.00
|
Это царство расположено в южном полушарии, причем в относительно высоких широтах. Оно включает южную часть Южной Америки, Новую Зеландию, незначительные не покрытые льдами участки Антарктиды и многочисленные мелкие острова, находящиеся в южной части Атлантического, Индийского и Тихого океанов.
РАСТИТЕЛЬНЫЙ ПОКРОВ ЗЕМНОГО ШАРА
В естественном растительном покрове земного шара различают зональную, интразональную и экстразональную растительность. Зональной растительностью называют ту, которая занимает плакоры – хорошо дренированные водораздельные пространства с почвами среднего механического состава.
Интразональной растительностью называют такую, которая развивается в особых, нетипичных для данной зоны условиях среды.
Экстразональная растительность также связана с какими-то особыми условиями среды и также встречаются пятнами на общем фоне зональной растительности.
Таким образом, зональная растительность создает основной фон, а интра- и экстразональная обычно встречаются в виде вкраплений и связанна с какими-то особыми условиями среды.
Различие между поясами и зонами: зоны значительно шире, имеют гораздо большую протяженность в пространстве. Это обусловлено тем, что в горизонтальном направлении климат изменяется несравненно медленнее, чем в вертикальном.
Характер зональной растительности какой-либо территории определяется прежде всего тем количеством тепла и влаги, которые получают растения, и распределением их по сезонам года.
РАСТИТЕЛЬНОСТЬ ЭКВАТОРИАЛЬНО-ТРОПИЧЕСКОГО ПОЯСА
Это самый теплый климатический пояс Земли. Однако в отношении условий увлажнения здесь наблюдается большое разнообразие. Есть районы, где выпадает огромное количество осадков, измеряемое несколькими тысячами миллиметров в год. Но в некоторых районах осадков, напротив, очень мало -100-200 мм и даже еще меньше.
В пределах тропического пояса выделяют следующие основные зональные типы растительности:
1. вечнозеленые дождевые тропические леса.
2. полулистопадные дождевые тропические леса.
3. влажные листопадные (муссонные) леса.
4. сухие листопадные (саванновые) леса.
5. саванны.
6. колючелистные ксерофильные редколесья.
РАСТИТЕЛЬНОСТЬ СУБТРОПИЧЕСКОГО ПОЯСА
Субтропические районы земного шара располагаются двумя сравнительно не широкими полосами приблизительно между 30 и 45 градусов северной и южной широты. Их разделяет экваториально-тропический пояс. Четкой границы между тропиками и субтропиками нет.
Климат субтропических районов в общем менее теплый, чем в тропиках. Изменение температуры по месяцам на протяжении года довольно значительно. Лето жаркое, а зима более или менее прохладная.
Влажность климата в разных субтропических районов не одинакова.
Субтропические районы, где выпадает значительное количество осадков на протяжении всего года, получили название субтропиков.
Таким образом, в субтропических районов различают три типа климата. Каждому из них соответствует определенное зональная растительность. Главнейшие типы зональной растительности субтропиков следующие:
1. влажные вечнозеленые субтропические леса.
2. жестколистные вечнозеленые сухие леса и кустарниковые заросли.
3. полупустыни.
4. пустыни.
РАСТИТЕЛЬНОСТЬ УМЕРЕННОГО ПОЯСА
Климат умеренного пояса в общем более холодный, чем в субтропиках. Здесь меньше тепла и ниже среднегодовые температуры. Во многих районах, за исключением приморских значительно ниже и абсолютные минимумы температуры. Характерно, что зима довольно продолжительная и морозная, это период устойчивых холодов. Лето же, напротив, может быть не только достаточно теплым, но даже жарким.
Главнейшие растительные зоны умеренного пояса – хвойные леса, летнезеленые лиственные леса, степи, пустыни. Все эти зоны хорошо выражены в северном полушарии и занимают здесь обширные территории.
Рассмотрим растительные зоны умеренного пояса северного полушария, начиная с более южных районов, примыкающих к субтропикам.
1. летнезеленые лиственные леса.
2. степи.
3. пустыни.
4. хвойные леса.
РАСТИТЕЛЬНОСТЬ ХОЛОДНОГО ПОЯСА
В холодном климатическом поясе еще меньше тепла, чем в умеренном, и общие условия существования растений особенно неблагоприятны. Лето очень короткое и холодное, причем температура хотя в общем и положительная, но большей частью довольно низкая. Безморозного периода совершенно нет. В любой летний день может наступить заморозок и выпасть снег. Зима очень долгая и крайне суровая.
Флора холодного пояса более бедна по сравнению с флорой умеренного пояса. Здесь могут существовать только не многие растения, особенно холодостойкие и нетребовательные к теплу.
В пределах холодного климатического пояса обычно выделяют только две зоны растительности: зону тундр и зону полярных пустынь.
7.Конспекты по зоологии.
Абдурахманов Г.М. Основы зоологии и зоогеографии.
АРЕАЛ
ОБЩИЕ СВЕДЕНИЯ ОБ АРЕАЛЕ
Ареал — часть территории или акватории земного шара, на которой постоянно встречаются популяции определенного вида или другого таксона животных.
В зоогеографии главным объектом изучения является ареал вида, поскольку вид — основная и исходная таксономическая категория живых организмов. Наряду с этим полноправно изучение как надвидовых, так и подвидовых ареалов.
Однако рассматривать ареал лишь как площадь не совсем правильно. Точнее представлять его как трехмерную структуру, поскольку водные биотопы имеют не только горизонтальное, но и вертикальное протяжение, да и наземные животные, распространенные и в горах, и на равнине, располагаются не только на площади, но и в некоем объеме.
Ареалы формировались в результате взаимодействия процессов эволюции животных и изменений среды их обитания в различные геологические эпохи. Поэтому изучение ареалов играет важную роль при выяснении их происхождения и палеогеографической эволюции. Кроме того, при сравнении ареалов одних и тех же видов через небольшие отрезки времени (10—20 лет) выявляется современная тенденция к расширению или сокращению площади, занятой популяциями вида.
Таким образом, карты распространения определенных таксонов остаются важнейшими орудиями труда зоогеографов.
КАРТИРОВАНИЕ АРЕАЛОВ
Правильное представление об ареале становится возможным только после его картографического изображения. Существуют различные методы картирования ареалов.
1. Точечный метод: местонахождение каждого обнаруженного вида обозначается точкой на контурной карте.
С помощью данного метода в отличие от других можно точнее определить пункт сбора, поимки особей того или иного вида либо его регистрации. Он оказывается единственно приемлемым методом, когда картируется распространение редкого или малоизученного вида. Этот метод имеет и недостатки. Он малопоказателен, так как требует известной доли воображения, чтобы представить себе форму и площадь ареала.
2. Контурный метод: дает возможность картировать границы ареала. С этой целью на контурной карте крайние, т. е. расположенные на границах ареала, точки соединяют сплошной линией, придавая изображению замкнутую форму.
3. Сеточный метод, растровой или метод формальных квадратов: При этом методе поле контурной карты разбивают на квадраты со стороной 50 км (для специальных целей масштаб может быть другим). Квадратом может служить и поле градусной сетки со стороной 1/4, 1/16 и т. д. градуса. Если в пределах квадрата обнаружены особи изучаемого вида, он затушевывается или обозначается условным знаком. Прочие квадраты остаются без обозначений.
На растровой карте разными условными знаками могут быть выделены местонахождения, известные по старым литературным данным и по новым находкам, что позволяет судить, меняются ли площадь и форма ареала со временем.
ТИПОЛОГИЯ АРЕАЛОВ
Каждый вид имеет свой собственный ареал, и в природе практически невозможно найти два абсолютно одинаковых по площади, расположению или по форме ареала, за исключением мелких островов, где различные виды животных могут населить всю его территорию, и тогда их ареалы будут изображаться на карте одинаково. Тем не менее все разнообразие ареалов можно свести к какому-то ограниченному числу типов.
Для номенклатуры ареалов важно использовать 3 оси координат, или 3 составляющие ареала: широтную (распространение с севера на юг), долготную (с запада на восток) и высотную (вертикальную, сверху вниз).
Широтная составляющая ареала (зональное распространение) является его основным географическим признаком. Она определяется в первую очередь климатическими, особенно температурными, факторами, прямо или косвенно влияющими на распространение большинства видов.
Высотная составляющая ареала (вертикальная поясность) в определенной степени аналогична широтной, поскольку также определяется главным образом температурными градиентами. По характеру вертикального распространения можно различить альпийские и монтанные (горные) виды. Последние заселяют среднегорья. Выделяют и промежуточные между ними — субальпийские формы. Часто приходится сталкиваться с равнинно-горным расселением.
Долготная составляющая ареала определяется степенью удаленности от океана, обусловливающей характер климата. Границы ареалов могут быть связаны с континентальным или морским, аридным или гумидным климатом.
Комбинация составляющих ареала позволяет охарактеризовать тип распространения и создать описательные классификации ареалов.
В основу классификаций должно быть положено одно из основных свойств ареалов — их размеры.
Можно построить иерархическую систему ареалов — от узколокальных до глобальных. Различают узкие и широкие ареалы. Как правило, наземные животные чаще имеют узкие ареалы, морские же — широкие. Иногда вид ограничен в своем распространении локальным районом, естественной географической (или фаунистической) областью. Такие виды называются эндемичными для данной области или просто эндемиками.
Особенно узкие ареалы типичны для видов, распространение которых ограничено какими-нибудь непреодолимыми преградами. Это прежде всего островные либо пещерные формы, обитатели горных долин или, наоборот, верхних зон горных хребтов. Нередки весьма узкие ареалы у нелетающих видов насекомых. Например, жужелицы-брызгуны обитают на Кавказе в пределах одного-двух хребтов. Все узкие ареалы свойственны эндемикам.
Еще более широкие ареалы, так называемые поли- или мультирегиональные, включают в себя несколько (не менее 2) фаунистических областей, часто располагающихся на разных материках.
Наконец, различают космополитные, или всесветные, ареалы. Они характерны чаще всего для морских животных, таких как кашалот, дельфин, касатка. Из наземных животных космополитами стали определенные насекомые-синантропы.
Между систематическим рангом группы и размером ареала существует корреляция: чем выше ранг группы, тем большую площадь она занимает. Следовательно, ареал рода больше ареала вида, ареал семейства больше ареала рода и т. д. Классы, как правило, распространены всесветно. Главными причинами, обусловливающими специфичность ареалов, являются экологическая валентность видов, их способность к расселению (вагильность), филогенетический возраст и скорость видообразования. Совокупность этих признаков и определяет величину ареала вида или их группы.
Формы ареалов, т. е. их очертания, столь же разнообразны, как и величина. Факторы, определяющие форму и величину ареала, чаще всего одни и те же. Очертания многих ареалов совпадают с ландшафтными или вообще с физико-географическими рубежами.
Как показал Ю.И.Чернов (1975), наиболее точное совпадение ареала с конфигурацией зоны наблюдается в тех случаях, когда вид связан с зональным типом растительности.
В горных странах ареалы многих видов вытянуты вдоль хребтов и тоже укладываются в одну из вертикальных зон — лесную, субальпийскую и т. д. Объясняется это тем, что вдоль определенного склона хребта наблюдаются одинаковые условия, чаще климатические.
Вытягивание ареала, обусловленное экологическими факторами, может привести к образованию так называемых ленточных, или линейных, типов ареалов. Они свойственны многим околоводным или водным животным, особенно обитателям литорали. Аналогичную форму имеют ареалы пресноводных рыб.
Нередко встречаются и сопряженные ареалы. Их очертания определяются границами ареала другого вида. Это бывает в том случае, когда между видами существуют экологические взаимоотношения, которые обычно носят характер пищевой связи.
Монофаги встречаются только там, где есть кормовое растение. Однако ареалы последних больше, поскольку распространение животных зависит не только от наличия корма, но и от других факторов. Значительно реже наблюдается сопряжение ареалов паразита и хозяина. Паразиту часто требуется промежуточный хозяин, а то и два, и отсутствие одного из них, даже при наличии основного хозяина, препятствует распространению этого паразита.
Все рассмотренные выше ареалы, от узколокальных эндемичных до громадных, занимающих весь материк, относятся к сплошным ареалам.
Встречаются ареалы, разделенные на части, т. е. прерывистые, или дизъюнктивные. О них следует говорить в том случае, когда отдельные части ареала обособлены друг от друга в такой степени, что никакая связь между заселяющими их популяциями вида невозможна.
Ареал может прерываться по-разному.
В основном он сплошной, но встречаются обособленные маленькие участки его, которые принято называть островными местонахождениями. Ареалы подобного типа считаются расчлененными (фрагментированными). Они возникают и в силу естественных причин, но часто своим происхождением обязаны деятельности человека.
Части дизъюнктивного ареала могут располагаться на одном материке, типы их разнообразны, но общим является то, что в этом случае между частями ареала нет преград в виде морских проливов или участков океана. На разделяющей территории природные условия тоже часто не отличаются от таковых в местах обитания видов, поэтому дать удовлетворительное объяснение причинам подобных разрывов довольно трудно. Как правило, части дизъюнктивного ареала связаны с одной ландшафтной зоной. Широко известными примерами такого рода являются амфиевро-азиатские неморальные ареалы амфибий — жерлянок рода Bombina.
Различают также арктоальпийский и бореомонтанный типы дизъюнктивного ареала. В первом случае основная часть ареала (зональная) находится в тундре, другая — в альпийском поясе гор Европы.
Особый и нередко встречающийся тип дизъюнкций представляют горные разрывы ареалов, когда отдельные их части находятся в верхних поясах горных хребтов и отделены друг от друга более или менее широкими долинами и межгорными понижениями.
Материковые разрывы ареалов.
Причины материковых разрывов ареалов разнообразны. Чаще разъединение объясняется фрагментацией сплошного ареала, вызванной изменением климата, конкуренцией более приспособленных форм, геоморфологическими перестройками или, наконец, деятельностью человека. В любом из этих случаев можно допустить либо вымирание отдельных популяций, либо вытеснение их другими, более приспособленными к новым условиям видами, что иногда подтверждается палеонтологическими находками в области разрыва.
Горные разрывы ареалов возникли в результате сложных геоморфологических процессов, происходивших в определенных частях земного шара.
Особый тип дизъюнктивных ареалов составляют ареалы, у которых между их частями находятся морские пространства. Подобные океанические дизъюнкции ареалов характерны для надвидовых таксонов — родов, семейств, отрядов и т. д. В пределах Северного полушария к данному типу ареалов относятся циркумполизональные, циркумполярные, циркумбореальные и циркумтемператные (расположены по всей умеренной зоне), а также амфиатлантические ареалы.
Очень обычны формы европейско-североамериканского и азиатско-американского разрывов. В этих случаях между ареалами родственных наземных и пресноводных таксонов находятся водные преграды — Атлантический океан, Берингов пролив или даже северная часть Тихого океана.
Среди океанических разрывов замечательны случаи биополярного и амфибореального распространения соответствующие группы животных встречаются в приполярных или умеренно холодных частях Земли и отсутствуют в тропиках и субтропиках.
Причины многих океанических разрывов ареалов не установлены.
Предложено несколько теорий, объясняющих происхождение подобных ареалов (теории мостов суши, оттесненных реликтов и др.).
Ареалы близких видов располагаются рядом, а иногда могут накладываться друг на друга (перекрывание ареалов). Бывает, что ареал одного вида, как правило, небольшой величины, охвачен гораздо большим ареалом другого вида. Из-за различных требований, предъявляемых видами к окружающей среде, перекрывание их ареалов — явление нередкое. Оно получило название симпатрии.
При изучении ареалов подвидов. В данном случае ареалы очень часто прилегают друг к другу, но никогда не перекрываются. Каждый подвид населяет определенную область видового ареала (подвиды аллопатричны), и подвидевые ареалы замещают друг друга, или викарируют.
Величина ареалов и их границы находятся в тесной взаимосвязи. Если размеры ареалов очень часто зависят от экологической валентности населяющих их видов, то значение экологических факторов, определяющих границы ареалов, еще более велико. Эти границы нередко обусловлены определенной комбинацией факторов среды или даже их одним градиентом. Особое значение имеют температура, освещение, питание, характер субстрата. Для наземных животных существенную роль, кроме того, играет влажность воздуха, для водных - химизм воды, содержание кислорода и т. д.
Существует разделение границ на климатические, ландшафтные, биоценотические. Их можно назвать экологическими границами, поскольку они определяются целой совокупностью факторов.
Границы ареалов, происхождение которых обусловлено минувшими событиями и не оправдывается экологической обстановкой настоящего времени, называются историческими или реликтовыми.
Биоценотические границы определяются взаимоотношениями разных видов в сообществе. Эти взаимоотношения носят разный характер (симбиоз, комменсализм, конкуренция и т. п.). Биологическое соперничество — одна из причин стабильности границ ареалов двух конкурирующих видов.
Однако стабильность границ — явление относительное и временное, тем более в настоящий период существования животного мира, когда в процесс расширения и сокращения ареалов вмешивается человек.
РАССЕЛЕНИЕ ЖИВОТНЫХ
Согласно дарвиновской теории эволюции, каждый вид возникает в определенной части земной поверхности и его первоначальный ареал в зависимости от характера формообразовательного процесса может быть большим или меньшим. Но со временем он расширяется, что происходит в результате расселения организмов. Под расселением, по определению А. И.Толмачаева (1974), следует понимать динамический процесс распространения видов за пределы занимаемой ими территории.
Изменения ареалов прослеживаются по изменениям очертаний их границ. Редко очертания границ меняются в течение одного или двух столетий, поэтому изменения устанавливаются с достоверностью.
Предпосылками расселения животных являются колебания численности особей в популяциях. Эти колебания происходят вследствие изменений соотношения между рождаемостью и смертностью особей.
Темпы расширения ареалов различны у разных видов, и это связано как с особенностями среды обитания, так и с характером самих видов. Способность видов к расселению зависит от их структурных, физиологических, поведенческих, популяционных и других свойств и носит название вагильности. Расселение животных — многофазный процесс, предусматривающий преодоление различных преград и препятствий.
Различают вагильность активную (организмы расселяются благодаря самостоятельному передвижению — полет, плавание, бег), пассивную (расстояние преодолевается за счет переноса ветром, течениями, человеком и т. д.) и смешанную (сочетание активной и пассивной вагильности).
Преграды к расселению можно условно свести к физическим и экологическим. К первым для сухопутных животных относятся, прежде всего, водные пространства, для водных организмов — большие участки суши, разделяющие водные бассейны. Ко второму непосредственно относится деятельность человека.
Так называемые биологические преграды обусловлены экологическими связями между видами животных и животных и растений. Отсутствие необходимого кормового объекта для монофага, хозяина для паразита, подходящей жертвы для хищника создает препятствия для расселения или вообще делает его невозможным. Расселение ограничивается и конкуренцией видов, занимающих одну и ту же экологическую нишу. На этом, в частности, основано явление викариата (замещения), характерное для подвидов одного вида, совершенно сходных по своим требованиям.
Пространство и время также выполняют функцию преград. Расселяясь из центра происхождения по территории, где нет преград и условия однообразны, вид, тем не менее, занимает не все пригодное для жизни пространство. Это означает, что расселение не завершилось и само расстояние служит своеобразной преградой.
Вопрос о возникновении и уничтожении преград или, иначе говоря, об изменении условий расселения животных — один из основных вопросов зоогеографии.
Активное расселение животных особенно характерно для обитателей Мирового океана и суши. Среди жителей внутренних водоемов в силу того, что последние изолированы друг от друга, оно наблюдается редко и свойственно в основном летающим насекомым, личинки которых ведут строго водный образ жизни (водяные жуки, стрекозы, ручейники и др.).
Активное расселение основано на различных формах самостоятельного движения (полет, бег, ползание, плавание и т.д.).
Пассивное расселение животных или их яиц и личинок осуществляется при помощи разнообразных факторов переноса.
Весьма характерна пассивная вагильность для пресноводных животных, особенно беспозвоночных. По всей видимости, вся пресноводная фауна в определенной мере имеет приспособления к пассивному переселению. Иначе невозможно объяснить поразительное сходство видового состава разобщенных водоемов, поскольку водные животные не имеют специальных органов для передвижения на суше. Способы пассивного расселения здесь весьма разнообразны: передвижение при помощи речных течений, транспортировка на плавающих предметах, разнос ветром, перенос мелких организмов на лапах водоплавающих птиц и т. д. Расселение проточными водами носит название гидрохории, а ветром — анемохории.
Расселение при помощи других животных — биохория — бывает случайным и закономерным. Особый ее случай — форезия — прикрепление на длительное время одного организма к другому для смены местообитания. Расселение паразитов основано на активном передвижении их хозяев.
Особый вид пассивного расселения — антропохорное, в котором участвует человек. Развитие средств сообщения между отдаленными странами, часто расположенными на разных материках, перевозка различных грузов и товаров делают неизбежным непроизвольный завоз самых разнообразных животных.
Несмотря на то что многие виды животных обладают разнообразными способностями к расселению, этого все же недостаточно для успешного преодоления преград и последующей колонизации нового района. Переселившимся организмам нужен биотоп или экологическая ниша, сходная с той, которую они занимали прежде.
Выживание переселенцев зависит как от внешних факторов, так и от их собственных свойств. Если параметры новой среды мало чем отличаются от прежних, колонизация может быть успешной.
Следует добавить, что хозяйственная деятельность человека, связанная обычно с изменением ландшафта, нередко способствует вытеснению аборигенных форм и создает благоприятные условия для новых колонистов.
ЦЕНТРЫ РАСПРОСТРАНЕНИЯ И
ПРОИСХОЖДЕНИЯ ВИДОВ
Расселение животных предполагает наличие первичного ареала — сравнительно небольшого района, где происходит видообразование, в ходе которого новая форма, дивергировавшая от родительской, приобретает видовую самостоятельность. Впоследствии таким же образом возникают другие близкие формы, объединяемые в роды, последние, в свою очередь, составляют таксономические единицы большего ранга — трибы, семейства и т. д. Каждая из этих единиц имеет свой ареал, слагающийся из видовых ареалов. Ареал рода (трибы, семейства) не просто сумма видовых ареалов: ареалы отдельных видов каждого рода не только дополняют друг друга, но и в определенной степени перекрываются. В таком случае общие очертания ареала рода определяют территорию, на которой встречается хотя бы один из его представителей. Поэтому при изучении родовых ареалов устанавливают не только их пределы, но и размещение в них отдельных видов. Чаще всего распределение видов в границах родового ареала оказывается неравномерным, и даже есть участки, где встречается всего лишь один вид. Места концентрации большого количества видов получили название очагов видового разнообразия или центров распространения.
Выявить центры видового разнообразия несложно. Однако эти центры не могут одновременно считаться и центрами их происхождения, т. е. родиной данных групп. Обнаруживая скопление большинства видов рода, можно лишь предположить, что здесь его родина и что отсюда началось расселение принадлежащих ему видов. Однако, учитывая, что ареалы динамичны и что первичный ареал (т. е. центр возникновения) мог располагаться на территории, где уже не существует ни одного вида рассматриваемого рода, проблема однозначно решается только при наличии соответствующего палеонтологического материала. Но так как ископаемые данные могут быть скудными или могут отсутствовать, то приходится пользоваться косвенными сведениями.
ПОНЯТИЕ О ФАУНЕ
Фауна — исторически сложившаяся совокупность видов животных, обитающих в данной области и входящих во все ее биогеоценозы.
Фауна объединяет все виды животных той или иной области (района, местности), хотя они и входят в различные биоценозы. Вместе с тем нельзя причислять к местной фауне виды, сознательно завезенные человеком и содержащиеся в зоопарках, аквариумах и т.д. Виды же, завезенные случайно, а также преднамеренно, но одичавшие или существующие без помощи человека, необходимо учитывать в составе местной фауны.
Каждая фауна характеризуется определенными, свойственными только ей признаками, позволяющими сравнивать ее с другими фаунами. Самым важным признаком любой фауны является ее видовой состав.
Существенным признаком любой фауны оказывается экологическая природа составляющих ее видов.
Главнейшей особенностью фауны являются ее связи с соседними, а также с более отдаленными фаунами. Эти связи можно охарактеризовать показателями общности систематического состава фауны, обычно выражаемой в процентах.
Наряду с фаунистическими связями не менее существен учет еще одного важнейшего признака — степени самобытности фауны, выражающейся в наличии эндемичных видов или родов. Чем выше систематический ранг эндемиков, тем самобытнее фауна. Так, присутствие в составе фаун эндемичных отрядов и семейств отражает длительность развития фауны в условиях изоляции. В то же время наличие только эндемичных видов, а тем более подвидов, свидетельствует об относительной молодости фауны и прочных связях ее с другими фаунами.
СТРУКТУРА ФАУНЫ
Каждая фауна обладает определенной систематической структурой, иначе говоря, специфичным распределением видов между родами, семействами и более высокими систематическими единицами. Структура фауны познается не только через количественное соотношение различных систематических единиц. Серьезное значение имеет также географический ее анализ т. е. установление сходства и различий в распределении входящих в нее видов. Виды, отличающиеся сходным распространением, представляют географические элементы фауны. Соответственно характеру распространения эти элементы носят определенные названия: северные, южные, западные, восточные. Такие названия пригодны для географического анализа фаун лишь ограниченных территорий. Если же рассматриваются ареалы видов той или иной фауны в целом, то наименования географических элементов будут другими. В зоогеографической литературе широко применяются термины: европейско-сибирский, восточносибирский (ангарский), центрально-азиатский, бореальный и др. Термин «заносные», или «адентивные», элементы означает, что данные виды натурализовались благодаря заносу извне и не являются единой географической группой.
Таким образом, географический анализ фауны дает представление о типе распространения входящих в нее видов. Но для познания фауны этого мало. Необходимо выяснить вероятное происхождение видов, как они попали в состав фауны и как происхождение каждого из них сказывается на распространении. Ответы на данные вопросы дает исторический (или генетический) анализ фауны. Он базируется на изучении ареалов не только видов, но и родов. При проведении его требуется, прежде всего, решить вопрос, какие элементы фауны возникли в пределах изучаемой территории и какие попали в результате расселения из других центров. Первые получили название автохтонных элементов, вторые — аллохтонных.
СРАВНИТЕЛЬНЫЙ АНАЛИЗ ФАУН
Сравнительный анализ начинается с изучения списков видов локальных фаун. При этом обращается внимание на качественные отличия, которые присутствуют лишь в одной из сравниваемых фаун. Количество видов (или других таксонов), характерных только для одной локальной фауны, служит показателем ее специфики.
При сравнительном анализе большое значение имеют определение степени общности и различия между двумя фаунами. Более точно это можно установить с помощью количественных методов. Для математического выражения степени сходства фаун двух районов предложен ряд формул. Наиболее часто применяется формула Жаккара, по которой определяют коэффициент общности (К), выражающийся в процентах:
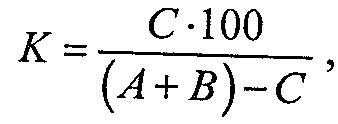
где С — число видов, общих для обеих фаун;
А, В — число видов соответственно в первой и второй фауне.
Используется также формула Сьеренсена:

Оба коэффициента в общем однотипно отражают степень сходства фаун сравниваемых районов.
ЭНДЕМИЗМ
Эндемичными систематическими единицами называются виды, роды и другие таксономические единицы, которые встречаются только в какой-то определенной области и нигде более. Эндемики представляют специфический компонент любой фауны и абсолютно отличают ее от других фаун.
Количество эндемиков в различных фаунах неодинаково. Наиболее высок процент эндемизма в островных фаунах, а на континентах — в районах с сильно расчлененным рельефом.
Для образования эндемичных видов требуется не только географическая изоляция, но и продолжительное время. Обилие эндемиков отражает длительность развития фаун. Различия между эндемичными и неэндемичными компонентами фауны не абсолютны. В большинстве фаун есть так называемые субэндемики, или почти эндемичные таксоны.
В историческом смысле можно говорить о прогрессивном и реликтовом эндемизме. Прогрессивными эндемиками (неоэндемиками) считаются виды или роды, возникшие в фауне недавно и еще не распространившиеся за пределы ее территории. К реликтовым эндемикам (палеоэндемикам) относятся животные, ареал которых в прошлом был гораздо больше, а в настоящее время они сохранились лишь на отдельной территории. Реликтовые эндемики занимают в фауне, как правило, изолированное положение.
ГЕНЕЗИС ФУНЫ
Каждая фауна имеет свою историю. Формирование фауны, или фауногенез, продолжается очень длительное время, измеряемое иногда целыми геологическими периодами. Описанием процесса зарождения и смены фаун занимается генетическая, или историческая, зоогеография. Лучшую опору для познания прежнего облика фауны дают ископаемые находки. При отсутствии палеонтологических материалов характер фауногенеза устанавливается по косвенным свидетельствам — данным исторической геологии, палеогеографии, выводам на основе теории эволюции. Большую пользу приносят также и собственные зоогеографические сведения о дизъюнкциях ареалов, эндемиках и реликтах.
ОСТРОВНЫЕ ФАУНЫ
Все разнообразие островов Мирового океана можно свести к нескольким основным типам. Главным критерием выделения типов является происхождение островов. Чаще всего они бывают океаническими и материковыми.
Остров материкового происхождения при отделении от материка наследует тот или иной (но не полный) набор местообитаний и связанных с ним видов.
На острове океанического происхождения первое время после его образования вообще нет никаких организмов. Современный органический мир таких островов формируется за счет видов, попавших на них путем залетов или заносов.
4 основных признака островных фаун: бедность и дефектность, выраженный эндемизм, ограниченное количество архаических форм и адаптивная радиация в рамках небольшого количества эндемичных групп.
Бедность и дефектность фауны — общий и характерный признак всех островов, проявляющийся в тем большей степени, чем остров меньше по площади и дальше находится от материка или другого острова.
Эндемизм — второй характерный признак островной фауны — является результатом изоляции острова от других участков суши.
Третья особенность островных фаун заключается в том, что они нередко являются последними убежищами архаичных видов животных.
Немногочисленные, а то и одиночные виды животных, попав на острова, оказываются родоначальниками процветающих и достаточно многочисленных эндемичных групп. Этот феномен адаптивной радиации в рамках небольшого количества эндемичных групп представляет собой четвертую особенность островных фаун.
ПРИНЦИПЫ И МЕТОДЫ
|
из
5.00
|
Обсуждение в статье: Голантарктическое царство |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы