 |
Главная |


Классификация нейронов
|
из
5.00
|
Тема 2. Структурно-функциональная характеристика нервной ткани. Биоэлектрические явления.
В основе современного представления о структуре и функции ЦНС лежит нейронная теория.
Нервная система построена из двух типов клеток: нервных и глиальных, причем число последних в 8 - 9 раз превышает число нервных. Однако, именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации.
Нейрон, нервная клетка, является структурно-функциональной единицей ЦНС. Отдельные нейроны, в отличие от других клеток организма, действующих изолированно, «работают» как единое целое. Их функции состоит в передаче информации (в форме сигналов) от одного участка нервной системы к другому, в обмене информацией между нервной системой и различными участками тела. При этом передающие и принимающие нейроны объединены в нервные сети и цепи.
В нервных клетках происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения.
Нейроны обладают рядом признаков, общих для всех клеток тела. Независимо от своего местонахождения и функций, любой нейрон, как всякая другая клетка, имеет плазматическую мембрану, определяющую границы индивидуальной клетки. Когда нейрон взаимодействует с другими нейронами, или улавливает изменения в локальной среде, он делает это с помощью мембраны и заключенных в ней молекулярных механизмов.
Нейроны, в отличие от других клеток, кроме тела (сомы) снабжены отростками. Многочисленные короткие древовидно разветвленные отростки - дендриты служат своеобразными входами нейрона, через которые сигналы поступают в нервную клетку. Они имеют шероховатую поверхность, создаваемую небольшими утолщениями - шипиками, словно бусинками, нанизанными на дендрит. Благодаря этому увеличивается поверхность нейрона и максимально повышается сбор информации.
Выходом нейрона является отходящий отросток - аксон, который передает нервные импульсы дальше другой нервной клетке или рабочему органу (Рис.1). Аксоны многих нейронов покрыты миелиновой оболочкой. Она образована швановскими клетками, многократно (до 10 и более слоев) «обернутыми» подобно изоляционной ленте вокруг ствола аксона. Однако, муфты швановских клеток, надетые на аксон, не соприкасаются друг с другом. Между ними остаются узкие щели - перехваты Ранвье. Только здесь нервное волокно непосредственно соприкасается с внеклеточной жидкостью. Поэтому, в нервной системе волна распространяющегося нервного импульса бежит не плавно, а движется скачками от одного перехвата к другому, что весьма ускоряет процесс распространения импульса.
Что же касается начальной части аксона в месте выхода его из тела клетки (область «аксонного холмика»), то она лишена миелиновой оболочки. Мембране этой немиелиновой части нейрона - так называемого начального сегмента обладает высокой возбудимостью. Поэтому ее называют пусковой зоной, так как именно отсюда начинается возбуждение нейрона.
Нет необходимости говорить о том, что даже для внутримозговых связей нужны очень длинные отростки, не говоря уже об аксонах, выходящих за пределы ЦНС - к мышцам, железам, внутренним органам. Собранные в пучки, они образуют нервы.
Если нейрон образует выходные связи с большим членом других клеток, то его аксон может многократно ветвиться, чтобы сигналы могли дойти до каждой из них, количество таких разветвлений колеблется от 1000 до 10000 и более. Кроме того, аксон способен отдавать дополнительные ветви - коллатерали, по которым возбуждение уходит далеко в сторону от магистрального пути.
Форма нервной клетки, ее размеры и расположение отростков разнообразны и зависят от функционального назначения нейрона.
Каждый отдельный нейрон уникален и неравноценен себе подобным, в отличие от других клеток организма. Величина нейронов весьма вариабельна: самые крупные в десятки и сотни раз больше самых мелких.
На вопрос, сколько же нейронов содержит мозг человека и высокоорганизованных животных никто точно ответить не может, но считается, что их количество измеряется приблизительно десятками миллиардов.
Классификация нейронов
Современная нейробиология предлагает два принципа классификации нейронов - по форме, строению и выполняемым ими функциям:
I. Классификация нейронов по форме: звездчатые, овальные, округлые, пирамидальные, клетки Беца (мозг).
II. Классификация нейронов по строению
1. Большинство нейронов состоят из тела, нескольких отходящих от него дендритов и одного аксона - мультиполярные нейроны:

Рис. Виды нервных клеток.
2. Нейроны, состоящие из тела, аксона и одного дендрита, называются биполярными.
| |
III. Нейроны делятся на возбуждающие и тормозные. Отсюда следует, что нейроны «специализирующиеся» на процессах возбуждения, называются возбуждающими, а на процессах торможения - тормозными.
IV. В зависимости от выполняемых функций обычно выделяют нейроны (рис ):
Афферентные (чувствительные, центростремительные, сенсорные) - передают импульсы (информацию) от рецепторов в ЦНС. Тела этих нейронов расположены вне ЦНС - в спинномозговых или черепно-мозговых ганглиях (рядом с головным и спинным мозгом). Афферентный нейрон имеет псевдоуниполярную форму, т.е. оба его отростка выходят из одного полюса клетки. Один из его отростков направляется на периферию, где заканчивается рецептором, а другой - в ЦНС. К афферентным нейронам также относятся нервные клетки, аксоны которых составляют восходящие пути головного и спинного мозга.
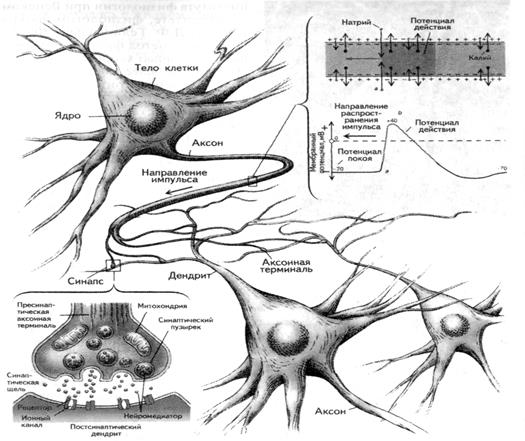
Рис Коммуникация нейрона
2. Эфферентные (эффекторные, двигательные и вегетативные) нейроныработают в центробежном режиме, т.е. они связаны с передачей нисходящих импульсов от вышерасположенных этажей нервной системы к нижерасположенным. Например, от коры к спинному мозгу, или от спинного мозга к рабочим органам. Для эфферентных нейронов характерна разветвленная сеть дендритов и один длинный аксон. Необходимо отметить, что количество эфферентных нейронов в 4-5 раз меньше афферентных.
3. Вставочные (промежуточные, интернейроны, сочетательные, ассоциативные) как правило, более мелкие клетки, осуществляющие связь между различными нейронами (в частности, афферентными и эфферентными). Они передают нервные импульсы в различных направлениях (горизонтальном, вертикальном) по ЦНС. Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов. В ЦНС преобладают промежуточные нейроны. Особое место занимают модуляторные нейроны, которые самостоятельно не запускают каких-либо реакций, но могут изменять уровень активности нервных центров, модулируя, таким образом, их реактивность.
4. Секреторные нейроны вырабатывают различные гормоны, выделяющиеся в кровь и осуществляющие гуморальную регуляцию работы различных органов и систем (нейроны гипоталамуса и гипофиза).
Из цепи функционально специализированных нейронов строятся рефлекторные дуги: простые (двухнейронные, моносинаптические) и очень сложные (полисинаптические). Связь между нейронами осуществляется посредством синапсов, чаще всего - химических. Нейрон, передающий информацию через синапс, называется пресинаптическим; получающий информацию нейрон называется постсинаптическим.
Глиальные клетки: их разновидности и функции.Нейроны в нервной системе окружены опорными и вспомогательными клетками, которые называются глиальными. Количество глиальных клеток в ЦНС в 5-10 раз превышает количество нейронов.
Клетки нейроглии плотно окружают всю сосудистую капиллярную сеть в мозговой ткани. Свободной остается лишь незначительная часть поверхности сосудов (около 15%). Выросты глиальных клеток могут располагаться с одной стороны на нейроне, с другой - на кровеносных сосудах. Это указывает на их важное значение в передаче питательных веществ и кислорода из крови в нервную клетку. Доказано, что нейроглия активно участвует в функционировании нейрона. В случае необходимости, компенсируется доставка питательных веществ и кислорода к активно «работающим» нейронам.
Клеткам нейроглии иногда приписывают также функцию своеобразной гидродинамической подушки, предохраняющей чувствительные и нежные образования нейронов от различных физических воздействий.
В последнее время появились сообщения об участии этих клеток в условно рефлекторной деятельности мозга и механизмах памяти.
Таким образом, система «нейрон – нейроглия» постоянно находится в состоянии гибкого ритмически колеблющегося равновесия. Но в этих процессах, происходящих в мозге, безусловно господствуют нейроны, и, пользуясь своим положением, тянут из нейроглии все, что ей нужно.
Глиальные клетки бывают нескольких типов. Т
| |
| |
| |
| |
| |
Аналогичные клетки в периферической нервной системе называются шванновскими клетками (клетками-спутниками, клетками-сателлитами).
Олигодендроциты образуют отростки, которые покрывают и изолируют нервные клетки и волокна. Олигодендроциты заключают их в складки своей наружной мембраны. При этом мембрана отростков олигодендроцитов, сопровождающих нервные волокна А и В типа, как бы накручивается вокруг соответствующего фрагмента каждого аксона. В результате эти клетки покрывают своей цитоплазматической мембраной ствол аксона в несколько слоёв с небольшими межклеточными промежутками между ними, называемыми перехватами Ранвье. После накручивания своих отростков вокруг аксона олигодендроцит начинает формировать специализированную клеточную оболочку, включая в её состав в качестве основного структурного белка миелин. Образовавшийся многослойный мембранный комплекс называется миелиновой оболочкой. Миелин, таким образом, образован мембранными белками и липидами, которые обусловливают белый цвет участков нервной ткани, состоящих преимущественно из миелинизированных волокон (белое вещество головного и спинного мозга).
| |
Олигодендроциты и шванновские клетки, заключающие в своих складках тела нейронов и немиелинизированные нервные волокна (С тип), могут одновременно образовывать изолирующие «футляры» для нескольких нейронов и их отростков.

Рис. Формы глиальных клеток
2. Астроциты имеют звёздчатую форму (рис.3). Некоторые из них снабжены тонкими цитоплазматическими отростками (фибриллярные астроциты), а часть - плотными отростками (протоплазматические астроциты).
Находятся астроциты преимущественно в сером и белом веществе головного и спинного мозга. Отростки заполняют пространства между сосудистыми стенками и нейронами. Это поддерживающие и изолирующие клетки, обеспечивающие транспорт веществ из кровеносных капилляров к нервным клеткам.
Астроциты обеспечивают нейроны питательными веществами, поступающими по сосудам (трофическая функция) и одновременно участвуют в формировании гематоэнцефалического барьера (ГЭБ), препятствующего поступлению из крови вредных веществ (защитная функция).
| |
4. Микроглия представлена мелкими клетками с множеством отростков. Клетки микроглии выполняют в ЦНС фагоцитарную функцию, удаляя погибшие нервные и глиальные клетки, вирусы и бактерии. Выполняет роль барьера между веществом мозга и омывающей его спинномозговой жидкостью; регулирует секрецию и состав спинномозговой жидкости.
развиваются в процессе эмбриогенеза из нервной трубки. Сложное взаимное расположение нейронов и клеток нейроглии обусловливает вид нервной ткани на разрезе, когда участки серого вещества чередуются с участками белого вещества. В спинном мозгу серое вещество занимает центральную позицию и имеет вид бабочки. В мозжечке и в полушариях большого мозга серое вещество образует ядра (участки скоплений нейронов при относительно небольшом количестве глиальных клеток, отделённые друг от друга белым веществом) и кору мозжечка и больших полушарий. Серое вещество коры большого мозга состоит из 6 слоёв нервных клеток, а в коре мозжечка таких слоёв три.
Биоэлектрические потенциалы в возбудимых тканях.К концу ХIХ и в начале ХХ стало известно, что электрические потенциалы образуются во всех живых тканях, а возникновение и распространение возбуждения связано с изменением электрического заряда на поверхности клеточной мембраны и внутри клетки.
В 1902 году Ю.Бернштейном была предложена мембранная теория возникновения электрических потенциалов живых тканей. Суть ее – потенциалы возникают за счет разности зарядов внутри и снаружи клетки.
теории натрий-калиевого насоса.
Было установлено, что величина внутриклеточного электрического потенциала у нервных клеток и волокон в покое составляет 70 милливольт (мВ). Эту величину называют мембранным потенциалом покоя (МПП).
Природа поляризации клеточной мембраны сегодня в основном определена. Мембрана аксона как и других клеток, тонкая, но достаточно прочная. Состоит мембрана из липидов, белков, мукополисахаридов. Мембрана легко проницаема для жирорастворимых веществ, молекулы которых легко проникают через липидный матрикс.
Крупные водорастворимые молекулы, в том числе анионы органических кислот практически не проникают мембрану и покидают клетку лишь путем экзоцитоза.
В то же время в мембране нервного волокна существуют каналы, проницаемые для воды, для малых молекул водорастворимых веществ и для малых ионов. Особое значение имеют каналы, проницаемые для ионов Nа +, К+, Cl‾, С²+. В нервной мембране обнаружены специфические натриевые, калиевые, хлорные и кальциевые каналы, то есть каналы избирательно пропускающие названные ионы. Эти каналы могут быть открытыми или закрытыми, то есть обладают воротными механизмами. В покое практически все натриевые каналы мембраны клетки закрыты, а большинство калиевых каналов открыты.
Для возникновения МПП, т.е. мембранного потенциала покоя, важно отличие ионного состава плазмы клетки от ионного состава внешней среды. В цитоплазме меньше ионов Nа+ и больше К+ и несравненно больше органических анионов. Катионные различия объясняются результатами работы так называемого калиево-натриевого насоса мембраны, непрерывно откачивающего Nа+ из клетки в обмен на К+. Движение ионов через мембрану называется активным ионным транспортом (рис..)
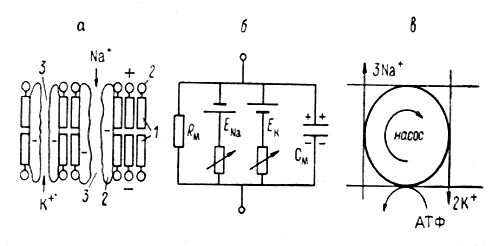
Рис.3. 1. Упрощенные схемы строения плазматической мембраны и ее механизмов, ответственных за формирование потенциала покоя и потенциала действия.
а - схема строения и б - эквивалентная электрическая схема мембран; в - схема работы ионного насоса. 1 - билипидный матрикс, 2- белковые молекулы; 3 - открытые катионные каналы.
Движение ионов, по сути, главный механизм формирования МПП. Но в образовании МПП участвует добавочный механизм – прямой электрогенный эффект натрий-калиевого насоса. Этот эффект зависит от поляризации мембраны, возникающей за счет неравенства числа ионов Nа+ и К+

Рис.3.2. Возникновение электрохимического равновесия на мембране.
Диффузионное давление (1) в равновесии с противоположной электростатической силой (2), стремящейся удержать вместе ионы с противоположными зарядами.
Разность зарядов между наружной и внутренней поверхностью клеточной мембраны в состоянии физиологического покоя клетки называется потенциалом покоя. В самой мембране МПП проявляется как электрическое поле значительной напряженности, которое обеспечивает закрытое и открытое состояние ворот натриевые каналов, обеспечивающих состояние покоя и готовности к возбуждению.
Потенциал действия или потенциал возбуждения нервных клеток возникает в ответ на достаточное по силе раздражение. Это очень быстрый, кратковременный электрический процесс, который проявляется, в первую очередь, изменением электрического состояния клеточной мембраны. Раздражение способствует перезарядке, реверсии, т.е. заряд мембраны уменьшается от нуля и приобретает отрицательное значение: внутренняя сторона мембраны заряжается положительно, наружная – отрицательно (рис.). Затем вновь восстанавливается первоначальный заряд. Колебания потенциала, возникающее в результате кратковременной перезарядке мембраны и последующего восстановления ее исходного заряда, называется потенциалом действия. В потенциале действия различают пик (длительность 0,5 – 0,8 мс) и следовые потенциалы (рис. )
Потенциал действия имеет несколько фаз:
1Фаза нарастания - потенциал действия начинается быстрым сдвигом в положительном направлении. В это время клеточная мембрана поляризована.
2. Овершут - деполяризация переходит за нулевую линию, и мембранный потенциал становится положительным.
3. Реполяризация - фаза, в течение которой восстанавливается исходный потенциал мембраны в покое. Последний участок фазы реполяризации для некоторых видов потенциала действия бывает замедлен.
4. Гиперполяризационный потенциал - кривая деполяризации быстро пересекает уровень потенциала покоя, так что на некоторое время потенциал становится более отрицательным, чем потенциал покоя.
5. Деполяризационным следовым потенциалом - примерно через 1 мс после начала потенциала действия наблюдается отчетливый перегиб кривой реполяризации.
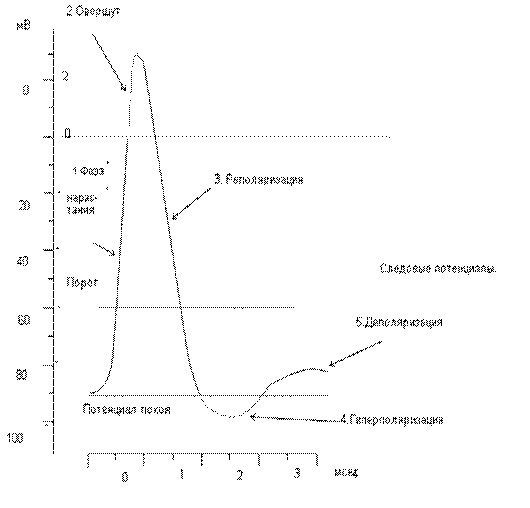 Рис. Фазы потенциала действия.
Рис. Фазы потенциала действия.
Современная мембранная теория объясняет механизм возникновения потенциала действия прохождением потоков ионов калия и натрия через каналы. Когда деполяризация достигает критической величины, за доли секунды, одновременно открываются максимальное количество натриевых каналов, проницаемость мембраны для ионов натрия увеличивается по сравнению с покоем в 500 раз. Ионы натрия устремляются внутрь клетки, унося с наружной поверхности мембраны положительный заряд, благодаря чему положительно заряженной становится внутренняя поверхность мембраны. Наружная же поверхность мембраны становится отрицательно заряженной.

Рис. Ионы натрия устремляются внутрь клетки, положительно заряжают внутреннюю поверхность мембраны. Наружная поверхность мембраны становится отрицательно заряженной.
Но такое положение длится всего 0,5 – 1,5 мс. Количество открытых натриевых каналов уменьшается, проницаемость для ионов натрия уменьшается до исходного уровня, этот процесс называется инактивацией механизма натриевой проницаемости. Но во время инактивации механизма натриевой проницаемости увеличивается проницаемость мембраны для ионов калия. Выходя из клетки, ионы калия выносят положительный заряд, чем восстанавливают исходную разность потенциалов и мембрана опять приобретает снаружи положительный заряд, изнутри – отрицательный.
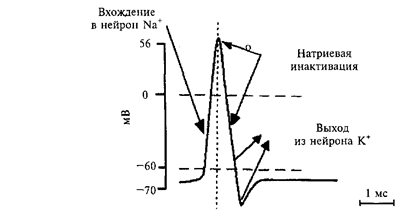
Рис. Схема событий при возбуждении нейрона.
Потенциал действия, возникнув в одном участке нервной клетки, нервного или мышечного волокна, быстро распространяется по всей ее поверхности. Потенциал действия по силе превышает в 5-10 раз силу раздражения, способную вызвать возбуждение, это необходимо для возбуждения участка клетки, находящегося в покое. Между возбужденным участком и находящимися в состоянии покоя, возникает разность потенциалов, приводящая к возникновению круговых токов.
Таким образом, потенциал покоя определяется ионами калия, а потенциал действия зависит от ионов натрия.
Скорость проведения возбуждения в волокнах скелетных мышц достигает 12-15 м/с, гладких мышц – 2-15, в безмякотных нервных волокнах – 0,5 – 3, в мякотных – 70-120 м/с.
Основные характеристики нервных тканей.Для любой живой клетки характерны следующие свойства: раздражимость, возбудимость, функциональная подвижность (лабильность).
Ι. Раздражимость – общее свойство любой живой ткани, клетки реагировать на раздражение изменением обмена веществ и энергии. Следует учитывать, что изменения носят местный характер, т.е. реагирует только участок ткани, который подвергается раздражению.
Раздражители можно условно разделить на три группы:
1. Физические механические, температурные, электрические, световые, невидимые глазом инфракрасное и ультрафиолетовое излучения, радиоактивное излучение.
2.Химические – кислоты, щелочи, яды.
3.Биологические – животные, растения, микробы и вирусы, паразиты высших млекопитающих
Кроме того, раздражители можно разделить на внешние (различные формы раздражения окружающей организм материи) и внутренние (изменение химического состава крови, лимфы, тканевой, спинномозговой жидкости).
Раздражители отличаются также и по силе воздействия.
Естественные раздражители, действующие на данную ткань в природных условия, называются адекватными. Изменения внешней или внутренней среды, являющиеся раздражителями, к восприятию которых не готовы все рецепторы или какой-либо, называются неадекватными. К последней группе относятся физические, механические, электрические раздражители.
ΙΙ. Возбудимость – это свойство нервной клетки отвечать на действие раздражителей изменениями ионной проницаемости мембраны и генерировать потенциал действия.
Минимальная сила (энергия) раздражителя, необходимая для раздражения клетки называется пороговой (порогом). Раздражители, сила которых меньше пороговой, называются подпороговыми. Раздражители, более сильные, чем пороговые – сверхпороговые. Возникнув в точке действия раздражителя, возбуждение способно охватывать всю клетку. Это свойство важно для нервной системы, особенно для ее проводящих путей. При отсутствии достаточных раздражителей в клетке наблюдается состояние физиологического покоя при условии текущего метаболизма. Реакции клетки под воздействием внешних факторов или других клеток организма, выражающиеся в снижении метаболизма, роста, возбудимости, называются торможением.
Для возникновения состояния возбуждения необходимы определенные условия:
а) сила действующего раздражителя должна быть не ниже пороговой;
б) достаточное время действия раздражителя (полезное время). Чем сильнее раздражитель, тем меньше времени потребуется для возбуждения ткани.
в) действие раздражителя с большой силой нарастания.
Изменение возбудимости нервной ткани при возбуждении. Для возбудимости тканей характерны фазовые изменения в зависимости от возникновения и развития импульсов. Во время деполяризации мембраны, при действии возбудителя, и появлении местного возбуждения происходит кратковременное повышение возбудимости. Когда местное возбуждение переходит в распространяющееся, возникает потенциал действия, а возбудимость ткани резко снижается. Возникает состояние, когда ткань после раздражения временно не реагирует на повторное раздражение любой силы – абсолютная рефрактерность (совпадает с пиком потенциала действия) (рис ).
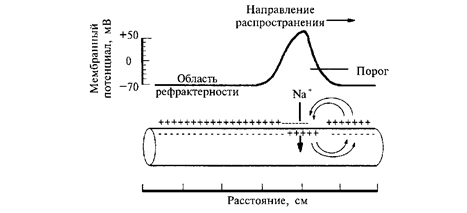
Рис. Распространение возбуждения по немиелинизированному нервному волокну: после прохождения потенциала действия в нервном волокне возникает зона невозбудимости, или рефрактерности.
Период восстановления ткани после абсолютной рефрактерности, когда возбудимость еще понижена называется относительной рефрактерности (совпадает с фазой реполяризации и переходом его в следовую поляризацию).
Затем наступает следующий период – фаза экзальтации – период повышенной возбудимости, совпадающий с периодом следовой поляризации. Со следовой гиперполяризацией совпадает фаза субнормальности, во время которой возбудимость ткани несколько снижена по сравнению с величиной возбудимости в состоянии физиологического покоя.
Очевидно, что фазы изменения возбудимости определены развитием и изменением потенциала действия в живой ткани. Изменение возбудимости зависит от действия натрий-калиевого насоса.
III. Важнейшим физиологическим свойством нервной ткани является функциональная подвижность или лабильность - скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Чем выше уровень процессов в тканях, тем быстрее процессы, составляющие отдельный импульс возбуждения и, следовательно, тем больше отдельных импульсов может образоваться в ткани за определенный отрезок времени. Максимальное число импульсов возбуждения, возникающее за 1 сек. в ответ на такое же число раздражений, называется мерой лабильности. Высокая лабильность характерна для мякотных соматических нервов, меньшая – для безмякотных нервов.
Самый частый ритм раздражений, на которые возбудимая ткань отвечает таким же частым ритмом возбуждения, отражает функциональное состояние ткани или лабильность. Лабильность может изменяться под действием разных условий, в частности зависеть от обмена веществ в ткани. Понижение и повышение физиологической лабильности в связи с деятельностью, проявляющееся более высоким или низким ритмом возбуждения названо усвоением ритма.
Наиболее выгодные в рабочем отношении частоты ритмов раздражения называют оптимум. При этом ритме раздражений новый импульс возникает в момент фазы экзальтации предшествующего импульса, в результате мышца сокращается максимально.
Пессимум ритма раздражения возникает, когда частота раздражений настолько высока, что превышает меру лабильности. Это вызвано тем, что раздражение действует в фазу абсолютной рефрактерности предыдущего импульса. Фаза рефрактерности может углубиться, лабильность будет продолжать снижаться и наступит торможение. Перечисленные явления характерны не только для двигательных реакций, но и для условно-рефлекторных, для секреторных реакций, деятельности сердечно-сосудистой системы.
Парабиоз, его стадии. Каким образом возбуждение переходит в торможение Н.Е.Введенский изложил в классическом труде «Возбуждение, торможение и наркоз».
При действии различных веществ, отличающихся по силе и частоте раздражения, мышца начинает отвечать одинаковыми сокращениями. Эту стадию Введенский назвал уравнительной или трансформирующей.
Следующая стадия сопровождалась сильными сокращениями мышцы при слабых или редких раздражениях, а при действии сильных раздражителей, наоборот, не сокращалась или почти не реагировала. Особенности стадии послужили тому, что ее назвали парадоксальной.
Третья стадия функциональных сдвигов сопровождалась состоянием, когда при воздействии на нерв раздражителя любой силы и частоты – мышца не сокращалась, развилось торможение. Стадия торможения заканчивается состоянием, при котором возбудимость и проводимость, как проявления жизни, отсутствуют, была названа Введенским парабиозом, а последовательные функциональные изменения - стадиями парабиотического процесса. При удалении раздражающих веществ с нерва парабиоз прекращается, функциональные свойства участка альтернирующей ткани восстанавливаются, проходя те же стадии в обратном порядке. Если воздействие раздражителей длительное, нерв отмирает.
|
из
5.00
|
Обсуждение в статье: Классификация нейронов |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы