 |
Главная |


Этапы уницикации энергетических субстратов в процессах катаболизма
|
из
5.00
|
Бред!
Субстрат - соединение, для которого данный фермент является специфическим катализатором;
Совокупность всех биохим. реакций, направленных на поддержание жизненных функций организма (рост, жизнедеятельность, воспроизведение и т.п.) принято называть обменом веществ и энергии или метаболизмом, различные продукты таких реакций – метаболитами. Последовательности биохимических реакций, из которых состоит обмен веществ называют метаболическими путями.
Обмен веществ в целом складывается из двух противоположных типов реакций – катаболических (катаболизм) и анаболических (анаболизм), протекающих в клетках одновременно. Катаболизм (диссимиляция) – это ферментативное расщепление крупных молекул (белков, жиров, углеводов и др.), осуществляемое преимущественно посредством реакцией гидролиза и окисления. В ходе катаболизма крупные органические молекулы расщепляются до простых веществ, что сопровождается выделением свободной энергии, которая запасается в форме, главным образом, энергии фосфатных связей АТФ.
В процессе катаболизма происходит постепенный и многоступенчатый распад сложных органических соединений, который можно разделить на три стадии.
На первой стадии распада крупные молекулы органических соединений расщепляются на свои составные части: белки – на аминокислоты, липиды – на глицерол, жирные кислоты и другие компоненты, полисахариды – на моносахариды, нуклеиновые кислоты – на нуклеотиды.
 На второй стадии продукты, образовавшиеся на первой стадии, путем анаэробного (без доступа кислорода) окисления превращаются в небольшое число более простых молекул. Глицерин, жирные кислоты, моносахариды и многие аминокислоты расщепляются до ацетил – КоА .
На второй стадии продукты, образовавшиеся на первой стадии, путем анаэробного (без доступа кислорода) окисления превращаются в небольшое число более простых молекул. Глицерин, жирные кислоты, моносахариды и многие аминокислоты расщепляются до ацетил – КоА .
Остальные аминокислоты при расщеплении образуют следующие соединения: α–кетоглутаровую кислоту (α–кетоглуторат), фумаровую кислоту (фумарат), щавелевоуксусную кислоту (оксалоацетат), янтарную кислоту (сукцинат).
На третьей стадии продукты, образовавшиеся на второй стадии, аэробно (с участием кислорода) окисляются через цикл ди- и трикарбоновых кислот (цикл Кребса, цикл лимонной кислоты), сопряженный с дыхательной цепью ферментов (электронно – транспортная цепь) до CO2 и H2O.
Освобождение химической энергии на разных стадиях катаболизма происходит в неодинаковой степени. На первой стадии катаболизма освобождается менее 1% химической энергии, на второй стадии – около 1/3, а на третьей – около 2/3 общего количества химической энергии, заключенной в органических соединениях. Около 40-50 % энергии, выделившейся при реакциях второй и третьей стадии катаболизма, рассеивается в виде тепла. Остальные 50-60 % запасаются в организме в форме энергии, главным образом, фосфатных связей АТФ; эта энергия в дальнейшем будет использована для реакций анаболизма и других нужд организма.
30. Энергетическая сущность явления окислительного фосфорилирования. Коэффициент фосфорилирования (Р/О).(см. вопрос 32)
Ок-е фосф-е, синтез АТФ из аденозиндифосфата и неорг. фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении орг. в-в в процессе клеточного дыхания. В общем виде О.ф. и его место в обмене в-в можно представить схемой:
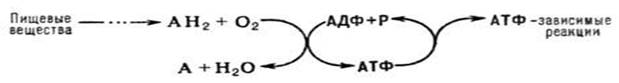
АН2 - орг. в-ва, окисляемые в дыхат. цепи (т. наз. субстраты окисления, или дыхания), АДФ - аденозиндифосфат, Р - неорг. фосфат.
Поскольку АТФ необходим для осуществления мн. процессов, требующих затраты энергии (биосинтез, совершение мех. работы, транспорт в-в и др.), ок. фосф. играет важнейшую роль в жизнедеятельности аэробных организмов. Образование АТФ в клетке происходит также благодаря др. процессам, напр. в ходе гликолиза и разл. типов брожения, протекающих без участия кислорода. Их вклад в синтез АТФ в условиях аэробного дыхания составляет незначит. часть от вклада ок. фосф. (ок. 5%).
У животных, растений и грибов протекает в митохондриях. У бактерий ферментные системы, осуществляющие этот процесс, находятся в клеточной мембране.
Механизм О.ф. можно представить схемой: Перенос электронов (дыхание)  А ~ В
А ~ В  АТФ А ~ В-высокоэнергетич. интермедиат. Наиб. признанием пользуется хемиосмотич. концепция сопряжения, предложенная в 1961 П. Митчеллом (Нобелевская премия). Согласно этой теории, своб. энергия транспорта электронов в дыхат. цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов Н+ (рис. 2, процесс 1). В результате на мембране возникает разность электрич. потенциалов
АТФ А ~ В-высокоэнергетич. интермедиат. Наиб. признанием пользуется хемиосмотич. концепция сопряжения, предложенная в 1961 П. Митчеллом (Нобелевская премия). Согласно этой теории, своб. энергия транспорта электронов в дыхат. цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов Н+ (рис. 2, процесс 1). В результате на мембране возникает разность электрич. потенциалов  и разность хим. активностей ионов Н+
и разность хим. активностей ионов Н+  (внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохим. потенциалов ионов водорода
(внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохим. потенциалов ионов водорода  между матриксом митохондрий и внеш. водной фазой, разделенными мембраной:
между матриксом митохондрий и внеш. водной фазой, разделенными мембраной: 
где R-универсальная газовая постоянная, T-абс. т-ра, F- число Фарадея. Величина  обычно составляет ок. 0,25 В, причем осн. часть (0,15-0,20 В) представлена электрич. составляющей
обычно составляет ок. 0,25 В, причем осн. часть (0,15-0,20 В) представлена электрич. составляющей  . Энергия
. Энергия 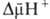 , выделяющаяся при движении протонов внутрь митохондрий по электрич. полю в сторону меньшей их концентрации (рис. 2, процесс 2), используется АТФ-синтетазой для синтеза АТФ. Т. обр., схему О.ф., согласно этой концепции, можно представить в след. виде:
, выделяющаяся при движении протонов внутрь митохондрий по электрич. полю в сторону меньшей их концентрации (рис. 2, процесс 2), используется АТФ-синтетазой для синтеза АТФ. Т. обр., схему О.ф., согласно этой концепции, можно представить в след. виде:
Перенос электронов (дыхание) 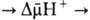 АТФ
АТФ
Сопряжение окисления и фосфорилирования через 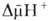 позволяет объяснить, почему О.ф., в отличие от гликолитич. ("субстратного") фосфорилирования, протекающего в р-ре, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрич. сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") О.ф. Энергия
позволяет объяснить, почему О.ф., в отличие от гликолитич. ("субстратного") фосфорилирования, протекающего в р-ре, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрич. сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") О.ф. Энергия  , помимо синтеза АТФ, может непосредственно использоваться клеткой для др. целей - транспорта метаболитов, движения (у бактерий), восстановления никотинамидных коферментов и др.
, помимо синтеза АТФ, может непосредственно использоваться клеткой для др. целей - транспорта метаболитов, движения (у бактерий), восстановления никотинамидных коферментов и др.
В дыхат. цепи имеется неск. участков, к-рые характеризуются значит. перепадом окислит.-восстановит. потенциала  и сопряжены с запасанием энергии (генерацией
и сопряжены с запасанием энергии (генерацией 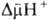 ). Таких участков, наз. пунктами или точками сопряжения, обычно три: НАДН: убихинон-редуктазное звено (
). Таких участков, наз. пунктами или точками сопряжения, обычно три: НАДН: убихинон-редуктазное звено (  0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (
0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (  ~ ~ 0,25 В) и цитохром-с-оксидазный комплекс (
~ ~ 0,25 В) и цитохром-с-оксидазный комплекс (  ~ 0,6 В)-пункты сопряжения 1, 2 и 3 соотв. (рис. 3). Каждый из пунктов сопряжения дыхат. цепи м.б. выделен измембраны в виде индивидуального ферментного комплекса, обладающего окислит.-восстановит. активностью. Такой комплекс,встроенный в фосфолипидную мембрану, способен функционировать как протонный насос.
~ 0,6 В)-пункты сопряжения 1, 2 и 3 соотв. (рис. 3). Каждый из пунктов сопряжения дыхат. цепи м.б. выделен измембраны в виде индивидуального ферментного комплекса, обладающего окислит.-восстановит. активностью. Такой комплекс,встроенный в фосфолипидную мембрану, способен функционировать как протонный насос.
Максимальная величина коэффициента фосфорили-рования, составляет 3 (т.к. в дыхательной цепи есть только 3 участка, где перенос электронов сопряжен с накоплением энергии, достаточным для образования АТФ ), если реакция окисления идет с участием НАД, и 2, если окисление субстрата протекает через флавиновые дегидрогеназы. Экспериментально определяемые значения Р/О, как правило, оказываются меньше 3. Это свидетельствует о том, что процесс дыхания не полностью сопряжен с фосфорилированием.
|
из
5.00
|
Обсуждение в статье: Этапы уницикации энергетических субстратов в процессах катаболизма |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы