 |
Главная |


Влияние углеводов на технологические свойства, качество и сохраняемость пищевых продуктов
|
из
5.00
|
Ни одно из основных пищевых веществ сырья и продовольственных товаров в такой степени не участвует в формировании ассортимента продовольственных товаров, как углеводы. Особенно велика их роль в бродильных производствах. Производство алкогольных изделий, пива, кваса, хлебобулочных изделий основано на способности углеводов к гидролизу и брожению. Практически за исключением питьевого молока и сливок огромный ассортимент молочных продуктов создан благодаря способности молочного сахара к брожению.
Велика роль углеводов в хлебопечении. Слово крахмал имеет немецкое происхождение и переводится как сильная мука (Kraft Mehl ). Cила крахмала заключается в его способности связывать большие количества воды (в несколько раз больше своей массы). Известно, что хлебопекарные свойства муки определяются газообразующей способностью муки и теста, а также его газоудерживающей способностью. При хорошем качестве дрожжей скорость образования в тесте углекислого газа зависит от количества сахара, имеющегося в муке и тесте. Однако того количества сахара, которое содержится в муке, недостаточно, чтобы процесс брожения теста шел достаточно долго. После расходования сахаров муки газообразующая способность определяется мальтозой, образующейся из крахмала под действием амилаз. Если мука имеет низкую амилолитическую активность, в тесте не появится в достаточном количестве мальтозы и глюкозы, брожение будет слабым и получится хлеб низкого качества, с недостаточно пористым, слабо разрыхленным, плотным мякишем.
Трудно переоценить роль реакций меланоидинообразования в формировании аромата и вкуса печеного хлеба. И в качестве обязательного исходного вещества в этих реакциях выступают углеводы.
Очень важно участие крахмала в процессах черствения хлеба. Черствение связано с так называемым процессом ретроградации крахмала, когда крахмал в условиях невысоких температур хранения хлеба теряет часть влаги. Эта влага остается в хлебе и повторный прогрев изделия может привести к связыванию выделившейся влаги и восстановлению свежести. Скорость ретроградации находится в зависимости от температуры клейстеризации крахмала зерна. Так, например, чем выше температура клейстеризации, тем меньше сроки хранения хлебобулочных изделий: Ржаной хлеб хранится более продолжительное время,чем пшеничный, так как крахмал пшеничной муки имеет более высокую температуру клейстеризации.
На технологические свойства ржи влияют также и слизистые вещества. Крахмал оказывает существенное влияние и на кулинарно-технологические свойства круп и макарон, в частности на увеличение объема при варке.
Крахмал из различных видов сырья сам по себе является самостоятельным сырьем для производства пищевых продуктов и изделий в кулинарии.
Гликоген (животный крахмал) оказывает существенное влияние на сроки хранения охлажденного мяса при хранении и косвенно на влагосвязывающую способность мяса и мясопродуктов. Это связано с тем, что при хранении при послеубойных изменениях часть гликогена превращается в молочную кислоту, которая, угнетая гнилостную микрофлору, продлевает сроки хранения мяса. Та же молочная кислота снижает рН мышечной ткани мяса до уровня изоэлектрической точки, что приводит белки в состояние окоченения и существенного снижения влагосвязывающей способности. Поэтому перед убоем животным создают условия, исключающие стресс для того, чтобы в полученном мясе содержалось достаточное количество гликогена.
Важным свойством сахарозы, имеющим большое практическое значение в отраслях пищевой промышленности, является карамелизация. При карамелизации образуются вещества различной степени конденсации, и соответственно, молекулярной массы и физических свойств. Самое низкомолекулярное – карамелан – имеет светло-коричневый цвет, карамелен характеризутся коричневой окраской потемнее. Наиболее темный из продуктов карамелизации – карамелин. Для карамелизации нужны очень высокие ( 160 и более градусов) температуры. В целом, карамелизация – сложный процесс, при котором образуются многие десятки различных веществ.
Одной из причин значительно меньших сроков хранения рыбы в охлажденном виде является меньшее содержание в ней гликогена.
Важная роль в формировании качества свежих плодов и овощей принадлежит пектиновым веществам. Протопектин укрепляет стенки плодов, делает их более устойчивыми при хранении, но малопригодными для потребления. Переход протопектина в пектин под влиянием ферментов переводит плоды из состояния съемной зрелости в потребительскую.
Характерным и важным свойством пектина является его способность давать студни в присутствии кислоты и сахара. Это свойство широко используется в кондитерской промышленности и общественном питании при производстве желе, джема, мармелада, пастилы и фруктовых карамельных начинок.
Содержание сахаров имеет существенное значение и для клубней картофеля. Накопление сахаров в значительных количествах делает картофель сладким и малопригодным для использования в общественном питании. Малопригоден такой картофель и для приготовления чипсов.
7. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ И БИОЭНЕРГЕТИКА
7.1. Митохондрии как внутриклеточные центры аэробного дыхания
Митохондрии — это особые органеллы клетки, внутриклеточные центры аэробного дыхания или «энергетические станции» клетки. Митохондрии обнаружены во всех клетках животных и растений, представляют собой овальные тельца размером примерно 0,5×З мкм. Форма митохондрий в разных клетках различна — от нитей до палочек, сфер и петель. Анализ серийных срезов одноклеточных организмов показывает, что митохондрии могут иметь гигантские размеры, почти равные самой клетке, неправильную разветвленную форму. Такие митохондрии обнаружены в клетках жгутиковых, в клетках мышц диафрагмы крысы, дрожжей.
Структура митохондрий изучена методом электронной микроскопии. В митохондриях различают две мембраны — наружную и внутреннюю, внутреннее пространство и межмембранное пространство. Внутреннее пространство заполнено так называемым матриксом — гелеобразной полужидкой массой. Внутренняя мембрана образует выпячивания, или кристы (рис. 18). Митохондриальные мембраны представляют собой липопротеиновые структуры толщиной 5—7 нм.
Наружная мембрана гладкая, хорошо проницаема для веществ, имеющих молекулярную массу менее 10 000. Для нее характерно высокое отношение количества фосфолипидов к белкам (примерно 1:1). В наружной мембране содержится ряд ферментов, участвующих в удлинении молекул насыщенных жирных кислот, а также белки, катализирующие окисление, не связанное с синтезом АТФ. Набор ферментов наружной мембраны различается в митохондриях разных организмов.

Рис. 16 Схема митохондрии. Л — в разрезе; Б — внутренняя мембрана: / — внутренняя мембрана, 2 — наружная мембрана, 3 — кристы, 4 — межмембранное пространство, 5 — матрикс
Внутренняя мембрана митохондрий покрыта со стороны матрикса грибовидными выростами, или «элементарными тельцами», которые в основном являются АТФазой. При обработке внутренней мембраны ультразвуком образуются «вывернутые частицы», у них грибовидные тельца находятся снаружи. Внутренняя мембрана непроницаема для большинства веществ, а также для Н+ и ОН-. Проникают только кислород и нейтральные молекулы с молекулярной массой меньше 150. Отношение фосфолипидов к белкам низкое, примерно 1:3. Внутренняя мембрана во всех клетках выполняет одинаковую функцию: сопряжение окисления с синтезом АТФ, т. е. является сопрягающей мембраной. Она содержит ферменты дыхательной цепи и синтеза АТФ.
В межмембранном пространстве митохондрий найдены ферменты аденилаткиназа и нуклеозиддифосфаткиназа.
У бактерий нет митохондрий, аэробное окисление и фосфорилирование протекают в цитоплазматической мембране и в особых мембранных образованиях — мезосомах. Мезосомы представлены двумя основными формами — ламеллярной и везикулярной, различающимися между собой укладкой мембраны. Хорошо выражены мезосомы везикулярного типа у представителей сем. Bacillaceae. Мезосомы ламеллярного типа чаще встречаются у кокков.
7.2. Пути использования энергии в клетке. Любое проявление жизнедеятельности, связаны с преобразованием энергии — движение, дыхание, воспроизведение, рост, реакции на возбуждение. Прямым источником энергии для большинства эндергонических процессов служит АТФ. Потенциальная химическая энергия АТФ используется для синтеза белков, жирных кислот, нуклеиновых кислот, стероидов и ряда других соединений. Биосинтетические реакции сопряжены с реакцией расщепления АТФ. Активирование обычно происходит за счет энергии, высвобождаемой при отщеплении фосфорильной группы от АТФ. Но для того чтобы потенциал последней мог быть использован для протекания эндергонического метаболического процесса, должен существовать механизм сопряжения, или гидролиз АТФ в клетке будет приводить просто к выделению тепла.
Разнообразные биосинтетические процессы в живом организме начинаются, с взаимодействия АТФ и исходного для биосинтеза соединения. Получив энергию одной из высокоэнергетических связей АТФ, это соединение активируется и становится способным вступать в биосинтез. Так активируются аминокислоты при биосинтезе белков, образуя предварительно аминоациладенилаты. Биосинтез полисахаридов также начинается с образования нуклеозиддифосфатсахаров при участии АТФ или УТФ.
7.3. Регуляторное теплообразование и свободное окисление. Оксигеназы и оксидазы. В организме среднего взрослого человека, весящего около 70 кг, каждые 24 ч генерируется и высвобождается 8000 кДж энергии. За это время не изменяются существенно ни масса тела, ни его структура и состав. Ися эта энергия, за исключением количества, которое потребовалось для выполнения физической работы, выделяется в виде тепла. Она необратимо рассеивается в окружающей среде. Этот процесс обеспечивает организм теплом, необходимым для поддержания температуры тела на уровне около 37°С.
Свободное окисление может протекать во внутренней мембране митохондрий, например, в случае воздействия особых веществ — разобщителей окисления и фосфорилирования.
В микросомах содержатся активные оксигеназы — ферменты, непосредственно присоединяющие кислород к различным субстратам. Различают две группы этих ферментов. Диоксигеназы катализируют присоединение обоих атомов молекулярного кислорода к окисляемому субстрату. В качестве простетической группы содержат гем или негемовое железо. Монооксигеназы (гидроксилазы) присоединяют только один атом О с образованием ОН-группы субстрата, другой атом восстанавливается при этом до воды с участием НАДН или НАДФН.
Микросомальное окисление — основная детоксицирующая система в организме животных и человека.
Кроме детоксицирующей функции оксигеназы у человека и животных играют определенную роль в некоторых реакциях биосинтеза (стероидных гормонов, желчных кислот, простагландинов) и других метаболических процессах (например, превращение циклических аминокислот).
В бактериальных и дрожжевых клетках оксигеназы способствуют поступлению и усвоению пластического материала, окисляя углеродсодержащие вещества и облегчая их поступление в клетки. У растений окисление органических соединений в монооксигеназных реакциях используется в основном для синтеза биорегуляторов.
Фенолоксидазы представляют собой медьпротеины, встречающиеся чаще в цитоплазме растительных клеток. Эти ферменты окисляют монофенолы в дифенолы, а остальные — до хинонов. Действием фенолоксидаз объясняется самопроизвольное потемнение срезов растительных тканей (картофель, фрукты, грибы), оно вызывается образованием хинонов с последующей спонтанной реакцией между хинонами.
Широко распространена в растениях аскорбатоксидаза — медьпротеин, также сосредоточенный главным образом в цитоплазме клеток. В растениях содержатся дыхательные цепи, в которых фенолоксидазы и аскорбатоксидаза функционируют как терминальные оксидазы. Эти окислительные системы локализуются вне митохондрий.
Регуляция энергетического обмена. Примером регуляции и саморегуляции энергетического обмена в организме является ускорение генерирования АТФ в работающей мышце. В покоящейся мышце, так же как и в других тканях, АТФ расходуется на поддержание постоянства внутренней среды и на непрерывно протекающие анаболические процессы. Но при выполнении механической работы потребность мышцы в энергии в форме АТФ может мгновенно возрастать в 20—200 раз. В это же время количество АТФ, содержащейся в мышечной ткани, невелико, достаточно не более чем для 0,5 с интенсивной работы. Большой вклад в энергетику мышцы вносит креатинфосфат, его в 3— 8 раз больше, чем АТФ; но запасы креатинфосфата также невелики, его расход должен возмещаться синтезом АТФ в процессах гликолиза и окислительного фосфорилирования. В период работы мышцы окислительное фосфорилирование усиливается, однако предельно возможное поглощение кислорода гораздо меньше, чем это требуется для получения достаточного количества энергии. Поэтому внезапное возрастание потребности в АТФ удовлетворяется в основном за счет гликолиза, скорость которого увеличивается в сотни раз. При этом используется запас мышечного гликогена, а также глюкоза, поступающая из крови. Молочная кислота, образующаяся при сокращении мышцы, диффундирует в кровь и ресинтезируется в печени в глюкозу и гликоген. АДФ и Фн, освобождающиеся при сокращении мышцы, используются в окислительном фосфорилировании и в генерирующих энергию реакциях гликолиза, повышая скорость этих процессов.
Другой пример регуляции энергетического обмена — эффект Пастера. Известно, что в анаэробных условиях дрожжи извлекают энергию из молекул глюкозы посредством брожения. Л. Пастер открыл, что в присутствии кислорода воздуха клетки дрожжей потребляют меньше глюкозы на единицу массы, чем в анаэробных условиях, при этом уменьшается накопление молочной кислоты. Торможение гликолиза в присутствии кислорода аэробным дыханием получило название эффекта Пастера. Он характерен для факультативно анаэробных клеток, включая клетки высших организмов.
Известно, что при недостатке АДФ скорость аэробного дыхания тормозится (дыхательный контроль). Таким способом регулируется скорость окислительного фосфорилирования. Д. Е. Аткинсон предложил характеризовать энергетическое состояние клетки количественно, вычисляя «заполнение» системы АТФ-АДФ-АМФ высокоэнергетическими фосфатными связями. Если все содержащиеся в клетке аденозинфосфаты находятся в форме АТФ, то система энергетически заполнена до предела, т. е. ее энергетический заряд равен 1,0. Если аденозинфосфаты находятся в форме АМФ, то система не содержит высокоэнергетических связей, она энергетически «пуста», ее заряд равен 0. Энергетический заряд системы АТФ-АДФ-АМФ вычисляется по уравнению
1 [АДФ] + 2 [АТФ]
Энергетический заряд = ─ ──────────────.
2 [АМФ] + [АДФ] + [АТФ]
От величины энергетического заряда зависят скорости реакций цикла трикарбоновых кислот и скорости других реакций, связанных с образованием или потреблением энергии. Например, у дрожжей уменьшение отношения АТФ/АМФ вызывает активацию цикла трикарбоновых кислот за счет активирования цитрат-синтазы и изоцитратдегидрогеназы. Важную регуляторную роль в тканях животных выполняет отношение АТФ/АДФ,
8. ЛИПИДЫ
8.1. Общая характеристика и классификация
Липидами называются неоднородные в химическом отношении вещества, общим свойством которых является хорошая растворимость в неполярных органических растворителях: эфире, ацетоне, хлороформе, четыреххлористом углероде, бензоле и т. п. В составе ряда липидов кроме этих компонентов встречаются фосфорная кислота, азотистые основания или углеводы. В экстракте, полученном при обработке животных или растительных тканей органическими растворителями, присутствуют обычно высшие и полициклические спирты, жирорастворимые витамины, которые некоторые авторы также относят к классу липидов.
Липиды могут быть классифицированы следующим образом: 1) нейтральные жиры и свободные жирные кислоты; 2) фосфолипиды; 3) гликолипиды; 4) стероиды; 5) воска; 6) терпены. Функции этого класса соединений важны и разнообразны.
Прежде всего, липиды в виде комплекса с белками являются структурными элементами мембран клеток и клеточных органелл. В связи с этим они определяют транспорт веществ в клетки. а так же участвуют в ряде других процессов, связанных с функционированием мембран.
Липиды служат также энергетическим материалом для организма. При окислении 1 г жира выделяется 39 кДж энергии. Одновременно липиды являются запасными веществами, в форме которых депонируется метаболическое топливо. Определенное исключение в этом отношении составляют бактерии: у большинства из них накопление энергии осуществляется в нелипидной форме (гликоген) и только у некоторых видов — в форме поли-3-гидроксимасляной.
В связи с хорошо выраженными термоизоляционными свойствами липиды сохраняют тепло в организме, особенно у морских и полярных животных, выполняя тем самым защитную функцию. В виде жировой прокладки липиды предохраняют тело и органы животных от механического повреждения, служат жировой смазкой для кожи. Восковой налет который находится на листьях и плодах растений защищает от избыточного испарения и проникновения микроорганизмов. Липидные компоненты бактерий определяют их чувствительность или резистентность к антибиотикам. Известны липиды которые имеют отношение к иммунитету (гликолипиды).
Регуляторной активностью обладают простагландины, полипреноловые коферменты — переносчики. От свойств и структуры мембранных липидов зависит активность мембраносвязанных ферментов, особенности протекания процессов окислительного фосфорилирования.
Есть мнение, что одна из наиболее важных функций липидов в мембранах заключается в их способности служить «кофактором твердого состояния», обеспечивая высокоупорядоченное, ориентированное, гидрофобное окружение для определенных ферментативных реакций. Являясь важнейшими компонентами нервных тканей, гликолипиды оказывают существенное влияние на функционирование нервной системы.
Важнейшее значение липиды имеют у бактерий, где они определяют таксономическую индивидуальность, дифференциацию видов, тип патогенеза и многие другие особенности.
8.2. Нейтральные жиры и жирные кислоты.
В связи с рекомендацией Международной номенклатурной комиссии нейтральные жиры принято называть триацилглицеролами. Это сложные эфиры трехатомного спирта глицерола и высших жирных кислот. Их общая формула
O
II
О СН2—О—С—R1
║ I
R2─C─O─CH O
I II
СН2—О—С—R3
R1, R2, R3 — остатки жирных кислот.
В молекуле глицерола могут быть этерифицированы как все три гидроксильные группы, так и две, и даже одна. Триацилглицеролы являются наиболее распространенной формой нейтральных жиров, хотя моно- и диацилглицеролы также встречаются в природе и играют важную роль в метаболизме липидов. Номенклатура нейтральных жиров основывается на названиях жирных кислот, входящих в их состав (например, тристеарин, олеодипальмитин).
Как запасные вещества триацилглицеролы обладают особыми преимуществами перед другими соединениями: углеводами, белками. Они не растворяются в воде и клеточном соке, не смешиваются с водой и поэтому не меняют физико-химические свойства цитоплазмы, до омыления ни в какие реакции в водной среде не вступают
Таблица 11. Природные жирные кислоты.
| Тривиальное название | Название по Женевской номенклатуре | Структура |
| Насыщенные | ||
| Лауриновая | н-Додекановая | СН3(СН2)10СООН |
| Миристиновая | н- Тетрад екановая | СН3 (СН2)12СООН |
| Пальмитиновая | н-Гексадекановая | СН3 (СН2)14СООН |
| Стеариновая | н-Октадекановая Ненасыщенные | СН3(СН2)16СООН |
| Пальмитоолеи новая | 9-Гексадеценовая | СН3 (СН2)5 СН=СН (СН2): COOH |
| Олеиновая | Цис-9-октадеценовая | СН3 (СН2)7-СН=СН (СН2)7 COOH |
| Линолевая | Цис-цис-9,12-октадека-диеновая | СН3 (CH2)4CH=CHCH2CH-CH (СН2)7 COOH |
| α-Линоленовая | 9, 12, 15-октадекатрие-новая | СН3СН2СН==СНСН2СН=СНСН2СН=СН (CH2)7 COOH |
| Арахидоновая | 5, 8, 11,1 4-Эйкозантет-раеновая | СН3 (СН2)4СН-СНСН2СН-СНСН2СН=СНСН2СН=СН(СН2)з COOH |
Жирные кислоты, входящие в состав липидов высших растений и животных, обладают рядом общих свойств. Почти все они содержат четное число атомов углерода, но чаще всего встречаются жирные кислоты с 16—18 атомами углерода, как насыщенные, так и ненасыщенные. Содержание ненасыщенных жирных кислот выше, чем насыщенных.
Так как ненасыщенные жирные кислоты имеют более низкую температуру плавления и содержащие их нейтральные жиры остаются жидкими даже при температуре ниже 5°С. Ненасыщенные жирные кислоты преобладают в растительных жирах (олеиновая, линолевая), называемых маслами.
В жирных кислотах растительного происхождения, содержащих более одной двойной связи, обнаружены сопряженные (конъюгированные) двойные связи: —СН = СН—СН = СН—, в то время как двойные связи в ненасыщенных жирных кислотах животных липидов обычно входят в дивинилметановую группировку —СН = СН— СН2—СН=СН—. Структурные свойства цепей жирных кислот имеют большое биологическое значение, особенно в мембранах.
Жирные кислоты, входящие в состав бактериальных липидов, характеризуются большим разнообразием. Установлено, что у большинства микроорганизмов длина цепи жирных кислот может достигать 26 углеродных атомов, но в основном это жирные кислоты, состоящие из 18 или меньшего числа углеродных атомов. Многие из насыщенных жирных кислот, от капроновой до стеариновой, в том числе и с нечетным числом атомов углерода, обнаруживаются почти у всех бактерий. Ненасыщенные жирные кислоты бактерий содержат чаще всего 16- или 18-углеродную молекулу с одной ненасыщенной связью. Ненасыщенные жирные кислоты с 5-7 двойными связями ( клупанадоновая, низиновая) содержатся в жирах морских животных и рыб. Некоторые из этих кислот обусловливают характерный запах рыбьих жиров. Они, как правило, относятся к так называемым омега-кислотам.
Жиры играют важную роль в метаболизме живых организмов. Из них получают мыло, олифу, масляные краски, основу для лекарственных мазей.
8.3. Фосфолипиды.
Молекула фосфолипидов образована остатками глицерина (или заменяющего его спирта сфингозина), жирных кислот, фосфорной кислоты, соединенной сложноэфирной связью с полярной группировкой (чаще азотсодержащей).
Фосфолипиды распространены в растительных и животных тканях, в микроорганизмах они являются преобладающей формой липидов. В отличие от нейтральных жиров фосфолипиды содержатся только в клеточных мембранах и очень редко в небольших количествах обнаруживаются в составе запасных отложений. Значительные количества фосфолипидов содержатся в сердце и печени животных, в семенах растений (например, в соевых бобах), в яйцах птиц. Особенно высоко содержание их в нервной ткани человека и позвоночных животных.
Фосфолипиды растворимы в органических растворителях, но избирательно: одни в этаноле, другие — в эфире, третьи — в бензоле. Большинство из них нерастворимо в ацетоне, это используют для отделения фосфолипидов от других липидов. Они нерастворимы в воде (но набухают); быстро окисляются на воздухе (из-за того что в молекуле есть ненасыщенные жирные кислоты), меняя при этом окраску от светло-желтой до коричневой.
Фосфолипиды легко образуют комплексы с белками, этим и объясняется преимущественное участие их в формировании клеточных оболочек и внутриклеточных мембран (составляют до 50% всех липидов биологических мембран). Благодаря особенностям химического строения (полярность молекулы) фосфолипиды обеспечивают одностороннюю проницаемость мембран. В соответствии с правилами Международной номенклатуры IUPAC класс фосфолипидов может быть разделен на глицеролфосфолипиды и сфингофосфолипиды.
Глицеролфосфолипиды. Общая схема строения глицеролфосфолипидов имеет следующий вид:
о
II
О СН2—О—С—R1
II I
R2-C-О—СН ОН
│ │
СН2—О —Р—О—X
║
О
X —полярная группа; R1 и R2 — радикалы жирных кислот.
В молекуле глицеролфосфолипидов лежит фосфатидная кислота, представляющая собой глицерин, у которого две спиртовые группы этерифицированы жирными кислотами, а одна, в 3-м положении, фосфорной кислотой. У большинства природных глицеролфосфолипидов насыщенные жирные кислоты (с 16—18 углеродными атомами) находятся в положении С-1.
В зависимости от строения полярной группы глицеролфосфолипиды делятся на:
1) фосфатидилхолины (старое название лецитины);
2) фосфатидилэтаноламины (старое название — кефалины);
3) фосфатидилсерины;
4) фосфатидилинозиты;
5) фосфатидилсахара;
6) фосфатидилглицерины;
7) фосфатидали, или плазмалогены.
В тканях животных и высших растений в больших количествах встречаются фосфатидилхолины и фосфатидилэтаноламины.
Сфингофосфолипиды. Липиды, относящиеся к этой группе, содержат те же компоненты, что и глицеролфосфолипиды (жирные кислоты, фосфат и Х-группу), только вместо глицерина они включают ненасыщенный аминоспирт сфингозин или его насыщенный аналог дигидросфингозин.
Н
│
Н3С─(СН2)12─ С = С─СНОН Н3С─(СН2)14─СНОН
│ │ I
Н Н - С—NH2 Н—С—NH2
I I
СН2ОН СН2ОН
Сфингозин (D-4-сфингенин) Дигидросфингозин (D-сфинганин)
Сфингофосфолипиды обнаружены в мембранах растительных и животных клеток. Их много в нервной ткани, в частности мозге, найдены и в составе липидов крови.
8.4. Гликолипиды. Гликолипиды содержат в составе молекулы углеводные остатки (чаще D-галактозы), фосфорная кислота в них отсутствует. Они находятся в основном в тканях мозга, есть также в клетках крови и других тканях; играют важную роль в функционировании биологических мембран. Представлены гликозилдиацилглицеринами и гликосфинголипидами.

| СН2ОН |

| (R.1 и R2 -радикалы жирных кислот) |
Гликозилдиацилглицерины. В их молекулу входит глицерин, этерифицированный двумя остатками жирных кислот, и одна или две молекулы моносахарида, связанных с глицерином β-гликозидной связью. В качестве углеводного компонента чаще всего встречается D-галактоза, реже D-глюкоза. Они составляют полярную голову липида. В отличие от фосфатидилсахаров в молекуле гликозилдиацилглицеринов нет фосфата. Представителями таких гликолипидов являются моногалактозилдиацилглицерин и дигалактозилдиацилглицерин. Они были выделены из листьев растений, где их концентрация приблизительно в 5 раз превышает концентрацию фосфолипидов из фотосинтезирующих бактерий.
Галактолипиды специфически связаны с хлоропластами. Ни один из перечисленных выше гликолипидов в животных тканях не обнаружен.
Гликосфинголипиды ганглиозиды, вовлечены в процесс приема сигналов, поступающих в клетки. Они активно участвуют в контроле и регуляции межклеточных контактов, рецепции пептидных гормонов, серотонина, некоторых вирусов и бактериальных токсинов. Структура ганглиозидов и их состав генетически контролируются гликозилтрансферазами, поэтому ганглиозиды обладают высокой тканевой специфичностью и выступают в роли антигенов клеточной поверхности.
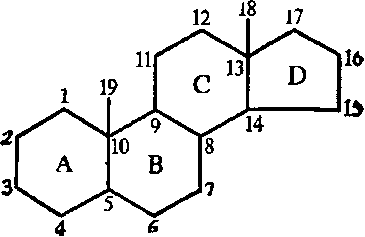
Стероиды. Стероиды — производные пергидроциклопентанфенантрена, имеющего следующую структуру:
Характерными особенностями почти всех природных стероидов является присутствие кислородсодержащего заместителя у С-3, наличие метильных групп, связанных с С-10 и С-13 и отсутствие двойных связей в циклах.
К стероидам относятся стеролы (или стерины), стериды, желчные кислоты, мужские и женские половые гормоны, гормоны надпочечников, витамины группы D, сердечные гликозиды, сапогенины, экдизоны, некоторые яды. В основе их классификации лежит структура заместителя, присоединяемого к 17-му углеродному атому ядра.
В клетках все стероиды присутствуют в следовых количествах, за исключением стеринов, содержание которых достаточно велико (до 2%).
8.5. Стерины (или стеролы)
К ним относятся стероиды, имеющие от 8 до 10 углеродных атомов в боковой цепи у С-17 и гидроксильную группу. Обнаружены в клетках животных (зоостерины), растений (фитостерины) и грибах (микостерины).
Одним из важнейших представителей зоостеринов является холестерин — одноатомный вторичный циклический спирт, относящийся к неомыляемым липидам. Он в значительном количестве содержится в липидах нервной ткани, где связан со структурными компонентами миелиновой оболочки, в липидах яиц и клеток спермы, в печени, надпочечниках кожном сале и в стенках эритроцитов.

В животных тканях холестерин находится как в свободйом состоянии, так и в виде сложного эфира с жирными кислотами — холестерида. Чаще всего в составе холестеридов обнаруживаются ненасыщенные жирные кислоты (олеиновая), а из насыщенных — пальмитиновая и стеариновая. В плазме крови только одна треть холестерина существует в виде свободного спирта, а две трети его этерифицировано жирными кислотами, переносчиком которых он и является. Образование эфиров происходит в стенках кишечника.
Холестерид ( R -радикал жирной кислоты)
Важнейшей биохимической функцией холестерина у позвоночных является его превращение в гормон прогестерон в плаценте, семенниках, желтом теле и надпочечниках, в результате чего открывается цепь биосинтеза стероидных половых гормонов и кортикостероидов. Другое направление метаболизма холестерина — образование желчных кислот и витамина D3. Кроме того, холестерин участвует в регулировании проницаемости клеточных мембран и предохраняет эритроциты крови от действия гемолитических ядов. Известно об активации холестерином ферментов цикла трикарбоновых кислот.
Холестерин преимущественно находится в животных тканях, некоторых водорослях и в очень небольшом количестве в пыльце и масле семян растений; из растительных продуктов выделен ряд фитостеролов. Весьма распространенной группой стеролов в растениях являются ситостерол С29Н49ОН и стигмастерол С29Н47ОН.
Особенно велико содержание стеролов в дрожжах и плесневых грибах. Основным стеролом в них является эргостерол С28Н43ОН.
Стериды. Стериды представляют собой эфиры стеринов (стеролов) и высших жирных кислот. Из жирных кислот в составе стеридов обнаружены в основном пальмитиновая, стеариновая и олеиновая. Однако в стеридах ланолина (восковидное вещество кожи и волос животных) обнаружены миристиновая, арахидоновая, церотиновая кислоты, а также ланопальмитиновая и ланостеариновая — специфические высшие жирные кислоты с разветвленной цепью.
Все стериды, как и стеролы,— это твердые бесцветные вещества. В природе, особенно у животных, они встречаются обычно в форме комплексов с белками.
8.6. Воска.
Воска представляют собой сложные эфиры высших моноатомных спиртов жирного (реже ароматического) ряда и высших жирных кислот. Помимо таких эфиров воска содержат некоторое количество свободных высших спиртов с четным числом атомов (от C22 до С32), свободных высших жирных кислот с очень длинной цепью (от C14 до C34), а также немного насыщенных углеводородов с нечетным числом углеродных атомов (от C21 до C37), душистых и красящих веществ. Общее количество этих примесей может достигать 50%. Обнаружены воска как у животных, так и у растений и даже у некоторых микроорганизмов.
Воска выполняют в организме в основном защитную функцию. Они образуют защитную смазку на коже, шерсти и перьях, покрывают листья, стебли, плоды, семена, а также кутикулу наружного скелета у многих насекомых. Восковой налет предохраняет от смачивания, высыхания и проникновения микробов. Из опытов видно, что удаление воскового слоя с поверхности плодов приводит к более быстрой их порче при хранении. Воска также являются главным липидным компонентом многих видов морского планктона — источника пищи для океанской фауны.
В состав восков входят как обычные жирные кислоты, содержащиеся в жирах,— пальмитиновая, стеариновая, олеиновая и др., так и жирные кислоты, характерные только для восков, — карнаубовая C24H48O2, церотиновая С27Н54О2 и др.
Среди высокомолекулярных спиртов, входящих в состав воска, наиболее изучены цетиловый спирт CH3(CH2)14CH2OH, мирициловый спирт С31Н63ОН, n-гексакозанол СНз(СН2)24СН20Н и др. Среди животных восков наибольшее значение имеют спермацет, ланолин и пчелиный воск.
Спермацет добывается из головы кашалота, где он находится в большом фиброзном мешке в углублении костей черепа и служит кашалоту звукопроводом при эхолокации, является эфиром цетилового спирта и пальмитиновой кислоты.
Спермацет используют в парфюмерии как основу при изготовлении кремов, мазей, так как он очень хорошо всасывается через кожу ( в настоящее время международными актами китобойный промысел запрещен, осуществляется ограниченно для научно-исследовательских целей), как и ланолин — смазочное вещество, покрывающее шерсть овец. По своему строению ланолин является стеридом. Он состоит из смеси эфиров двух стеринов — ланостерина и агностерина и жирных кислот ланолиновой, пальмитиновой, стеариновой и др.
Пчелиный воск вырабатывается специальными железами рабочих пчел. По своему химизму это преимущественно мирицилпальмитат, но в нем есть и другие сложные эфиры, а также свободные жирные кислоты, свободные высшие спирты (цериловый спирт и др.) и углеводороды. В его составе обнаруживаются вещества, обусловливающие цвет и запах, а также минеральные соединения.
Пчелиный воск находит применение в медицине и различных отраслях промышленности благодаря сочетанию пластичности с кислотоустойчивостью, водо- и электроизоляционными свойствами: литейной, кожевенной, автомобильной, авиационной, текстильной, электрической, радиотелефонной, пищевой, фармацевтической, стекольной, парфюмерной, гальванопластике и др.
8.7. Терпены.
Общая характеристика терпенов. В основе терпеновых соединений лежат изопреновые остатки, чаще всего соединенные между собой «голова к хвосту», но иногда возможно апериодическое расположение изопреновых единиц - «хвост к хвосту».
СН3
│
(Голова) Н2С=С—СН=СН2 (Хвост)
Изопрен
К терпенам относятся эфирные масла (лимонен, пинен, гераниол, камфора, ментол), смоляные кислоты и каучук, различные растительные пигменты (каротины, ликопин и др.), а также витамин А и сквален из животных тканей. Химически родственны терпенам витамин Е, витамин К, кофермент Q.
В зависимости от числа изопреновых группировок все терпены делятся на монотерпены (2 группировки), сесквитерпены (3), дитерпены (4), тритерпены (
|
из
5.00
|
Обсуждение в статье: Влияние углеводов на технологические свойства, качество и сохраняемость пищевых продуктов |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы