 |
Главная |


Анаэробный распад углеводов
|
из
5.00
|
Анаэробный распад углеводов обычно протекает в мышцах при выполнении интенсивных нагрузок. По своему содержанию анаэробный распад соответствует первому этапу ГДФ-пути. Однако из-за высокой скорости этого процесса образующиеся в больших количествах НАД-Н2 и пируват не полностью окисляются в митохондриях. В этом случае большая часть НАД-Н2 передает атомы водорода пировиноградной кислоте непосредственно в цитоплазме 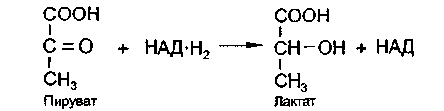
В этом случае все реакции протекают анаэробно(без участия митохондрий и потребления кислорода) и приводят к образованию и накоплению лактата (молочной кислоты). Такой анаэробный распад углеводов получил название анаэробный гликолиз,или просто гликолиз Гликолиз полностью протекает в цитоплазме клеток, и синтез АТФ происходит только анаэробно 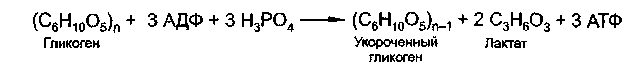
Анаэробный распад гликогена является дополнительным способом получения АТФ при выполнении интенсивной мышечной работы, протекающим без участия митохондрий и потребления кислорода. Анаэробный распад глюкозы наблюдается главным образом в красных клетках крови (эритроцитах), в которых отсутствуют митохондрии, и при этом образуется на 1 молекулу АТФ меньше.
Катаболизм жиров
Использование жира в качестве источника энергии начинается с его выхода из жировых депо в кровяное русло. Этот процесс называется мобилизациейжира. Мобилизация жира ускоряется под влиянием симпатической нервной системы и гормона адреналина. В печени жир прежде всего подвергается гидролизу и превращается, так же как и в кишечнике, в глицерин и жирные кислоты.
Образовавшийся глицеринлегко переходит в фосфоглицериновый альдегид,который является также промежуточным продуктом распада углеводов и поэтому вовлекается в углеводный обмен.
Жирные кислоты,являясь веществами химически неактивными, вначале активируются с использованием энергии АТФ и связываются со своим переносчиком - коферментом А(комплекс жирная кислота -кофермент Аназывается ацилкофермент А).
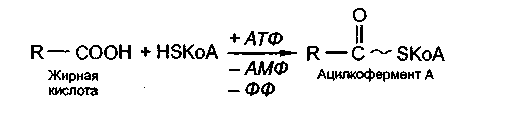
Образовавшийся ацилкофермент А поступает в митохондрии, где происходит окисление жирной кислоты.
На первом этапе, называемом р-окислением, от жирной кислоты, связанной с коферментом А, дважды отщепляется по два атома водорода, которые затем по дыхательной цепи передаются на молекулярный кислород. В итоге образуется вода, и за счет выделяющейся при этом энергии осуществляется синтез пяти молекул АТФ. Завершается р-окисление отщеплением от жирной кислоты фрагмента в виде ацетилкофермента А. Р-окисление многократно повторяется до тех пор, пока жирная кислота полностью не превратится в ацетил-КоА, количество молекул которого равно половине числа атомов углерода в исходной жирной кислоте. Какуже отмечалось, каждое отщепление ацетилкофермента А сопровождается синтезом пяти молекул АТФ, осуществляемым тканевым дыханием.
Вторым этапомокисления жирных кислот является цикл трикарбоновых кислот (ЦТК) или цикл Кребса, в котором происходит дальнейшее окисление остатка уксусной кислоты, входящей в ацетилко-фермент А, до углекислого газа и воды. При окислении одной молекулы ацетилкофермента А выделяется 12 молекул АТФ.
В целом окисление жирных кислот до С02 и Н20 дает большое количество энергии. Например, в случае окисления пальмитиновой кислоты(С15Н31СООН) семь раз протекает р-окисление, что приводит к образованию 35 молекул АТФ и 8 молекул ацетилкофермента А. При дальнейшем окислении 8 молекул ацетилкофермента А в цикле Кребса еще синтезируется 96 молекул АТФ. Вычтя из полученной суммы молекул АТФ одну молекулу, энергия которой была затрачена на активацию жирной кислоты, получаем окончательный результат: при окислении молекулы пальмитиновой кислоты образуется 130 молекул АТФ.
При избыточном образовании ацетил-кофермента А, что обычно бывает в печени, вместо цикла Кребса происходит реакция конденсации.В результате конденсации остатки уксусной кислоты, соединяясь попарно, превращаются в кетоновые тела, а кофермент А выделяется в свободном виде. С током крови кетоновые тела поступают во все ткани. Однако большая часть кетоновых тел извлекается из крови органами, имеющими высокие энергозатраты: миокардом, скелетными мышцами, почка-В этих органах с участием их собственного кофермента А кетоновые тела вновь переходят в ацетилкофермент А.Далее ацетилкофермент А, окисляясь в цикле Кребса, дает этим органам необходимую энергию для их функционирования. Особенно велика роль кетоновых тел при обеспечении энергией продолжительных физических нагрузок.
При накоплении кетоновых тел в крови возможно образование ацетона,который выводится легкими.
Синтез жиров
Синтезируются жиры из глицерина и жирных кислот. Глицерин в организме возникает при распаде жира (пищевого или собственного), а также легко образуется из углеводов. Жирные кислоты синтезируются из ацетилкофермента А - универсального метаболита организма. Для этого синтеза еще необходимы водород (в форме НАДФН2) и энергия АТФ. В организме синтезируются только насыщенные и мононенасыщенные (имеющие одну двойную связь) жирные кислоты. Кислоты, содержащие две и более двойных связей в своей молекуле (полиненасыщенные), в организме не синтезируются и должны поступать с пищей. Для синтеза жира также могут быть использованы жирные кислоты - продукты гидролиза пищевого и собственного жиров.
Все участники синтеза жира должны быть в активном виде: глицерин в форме глицерофосфата, а жирные кислоты в форме ацилко-фермента А. Синтез жира осуществляется в цитоплазме клеток (преимущественно жировой ткани, печени, тонкой кишки) и протекает по следующей схеме 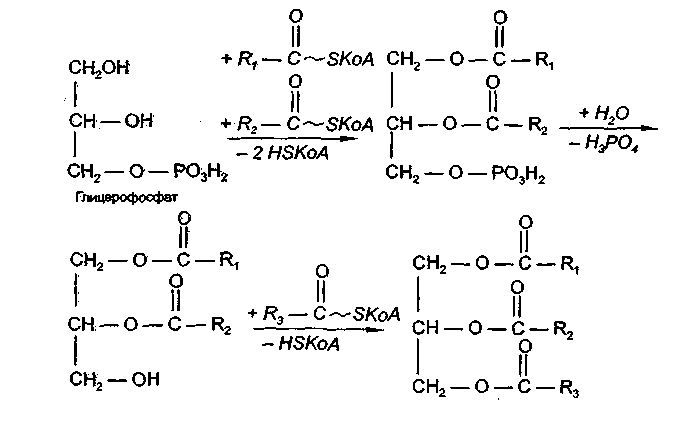
Следует подчеркнуть, что глицерин и жирные кислоты могут быть получены углеводов. Поэтому при избыточном потреблении углеводов на фоне малоподвижного образа жизни развивается ожирение.
Лекция 4. Обмен белков
Катаболизм белков
Белки, входящие в состав клеток организма, также подвергаются постоянному распаду под влиянием внутриклеточных протеолитических ферментов, называемых внутриклеточными протеиназамиили катепсинами.Эти ферменты локализованы в специальных внутриклеточных органоидах – лизосомах. Под действием катепсинов белки организма также превращаются в аминокислоты. (Важно отметить, что распад как пищевых, так и собственных белков организма приводит к образованию одних и тех же 20 видов аминокислот.) В сутки расщепляется примерно 200 г белков организма. Поэтому в течение суток в организме появляется около 300 г свободных аминокислот.
Синтез белков
Большая часть аминокислот используется для синтеза белков. Синтез белков происходит при обязательном участии нуклеиновых кислот.
Первый этап синтеза белка - транскрипция - осуществляется в клеточном ядре с использованием ДНК как источника генетической информации. Генетическая информация обусловливает порядок расположения аминокислот в полипептидных цепях синтезируемого белка. Эта информация закодирована последовательностью азотистых оснований в молекуле ДНК. Каждая аминокислота кодируется сочетанием трех азотистых оснований, называемым кодоном, или триплетом. Участок молекулы ДНК, содержащий информацию об определенном белке, получил название «ген».На этом участке ДНК во время транскрипции по принципу комплементарности синтезируется информационная РНК (иРНК). Эта нуклеиновая кислота представляет собой копию соответствующего гена. Образовавшаяся иРНК выходит из ядра и поступает в цитоплазму. Аналогичным образом на ДНК как на матрице происходит синтез рибосомных (рРНК) и транспортных (тРНК).
В ходе второго этапа – рекогниции (распознавания), протекающего в цитоплазме, аминокислоты избирательно связываются со своими переносчиками - транспортными РНК(тРНК). Молекула каждой тРНК представляет собой короткую полинуклеотидную цепь, содержащую примерно 80 нуклеотидов и частично закрученную в двойную спираль, что приводит к возникновению конфигурации «изогнутого клеверного листа». На одном конце полинуклеотидной цепи у всех тРНК находится нуклеотид, содержащий аденин. К этому концу молекулы тРНК присоединяется аминокислота. Петля, противоположная месту присоединения аминокислоты, содержит антикодон, состоящий из трех азотистых оснований и предназначенный для последующего связывания с комплементарным кодоном иРНК. Одна из боковых петель молекулы тРНК обеспечивает присоединение тРНК к ферменту, участвующему в рекогниции, а другая, боковая, петля необходима для присоединения тРНК к рибосоме на следующем этапе синтеза белка.
На этом этапе в качестве источника энергии используется молекула АТФ. В результате рекогниции образуется комплекс аминокислота-тРНК. В связи с этим второй этап синтеза белка называют активацией аминокислот.
Третий этап синтеза белка - трансляция - происходит на рибосомах. Каждая рибосома состоит из двух частей - большой и малой субчастиц. По химическому составу обе субчастицы состоят из рРНК и белков. Рибосомы способны легко распадаться на субчастицы, которые снова могут соединяться друг с другом, образуя рибосому. Трансляция начинается с диссоциации рибосомы на субчастицы, которые сразу же присоединяются к начальной части молекулы иРНК, поступающей из ядра. При этом между субчастицами остается пространство (так называемый туннель), где располагается небольшой участок иРНК. Затем к образовавшемуся комплексу рибосома - иРНК присоединяются тРНК, связанные с аминокислотами. Присоединение тРНК к этому комплексу происходит путем связывания одной из боковых петлей тРНК с рибосомой и связывания антикодона тРНК с комплементарным ему кодоном иРНК, находящимся в туннеле между субчастицами рибосомы. Одновременно к комплексу рибосома - иРНК могут присоединиться только две тРНК с аминокислотами.
Благодаря специфическому связыванию антикодонов тРНК с кодонами иРНК, к участку молекулы иРНК, находящемуся в туннеле, присоединяются только молекулы тех тРНК, у которых антикодоны комплементарны кодонам иРНК. Поэтому эти тРНК доставляют в рибосомы только строго определенные аминокислоты. Далее аминокислоты соединяются друг с другом пептидной связью и образуется дипептид, который связан с одной из тРНК. После этого рибосома передвигается вдоль иРНК ровно на один кодон (это перемещение рибосомы называется траислокацией).
В результате транслокации свободная (без аминокислоты) тРНК отщепляется от рибосомы, а в зоне туннеля появляется новый кодон, к которому присоединяется по принципу комплементарности еще одна тРНК с аминокислотой, соответствующей этому кодону. Доставленная аминокислота соединяется с ранее образовавшимся дипептидом, что приводит к удлинению пептидной цепи. Далее следуют новые транслокации, поступление на рибосому новых тРНК с аминокислотами и дальнейшее удлинение пептидной цепи.
Таким образом порядок включения аминокислот в синтезируемый белок определяется последовательностью кодонов в иРНК. Завершается синтез полипептидной цепи при поступлении в туннель особого кодона, который не кодирует аминокислоты и к которому не может присоединиться ни одна тРНК. Такие кодоны называются терминирующими.
В итоге за счет описанных трех этапов синтезируются полипептиды, т. е. формируется первичная структура белка. Высшие (пространственные) структуры (вторичная, третичная, четвертичная) возникают самопроизвольно.
Синтез белков - процесс энергоемкий. Для включения в молекулу синтезируемого белка только одной аминокислоты требуется не менее трех молекул АТФ.
|
из
5.00
|
Обсуждение в статье: Анаэробный распад углеводов |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы