 |
Главная |


Полимерноре взаимодействие генов
|
из
5.00
|
Полимерное взаимодействие генов
Полимерия – тип взаимодействия неаллельных генов, при котором несколько пар неаллельных генов действуют на развитие одного признака. Такие гены называются полимерными (от греч. полис – много и мерос – часть). Их обозначают одной буквой, но с разными индексами, которые указывают на число аллельных пар в генотипе, обусловливающих развитие конкретного признака (А1А1А2А2).
Различают два варианта полимерии: с суммирующим действием генов и без усиления генов друг другом.
• Суммирующее действие полимерных генов. В этом случае степень проявления признака зависит от числа доминантных аллелей в генотипе особи. Так, красная окраска зерен пшеницы определяется двумя и более парами генов. Каждый из доминантных генов этих аллелей определяет красную окраску, рецессивные гены определяют белый цвет зерен. Один доминантный аллель дает не очень сильно окрашенные зерна. Если в генотипе присутствуют два доминантных аллеля, интенсивность окраски повышается. Лишь в том случае, когда организм оказывается гомозиготным по всем парам рецессивных генов, зерна не окрашены. Таким образом, при скрещивании дигибридов происходит расщепление в отношении 15 окрашенных к одному белому. Но из 15 окрашенных один будет иметь интенсивный красный цвет, т.к. содержит четыре доминантных аллеля, четыре будут окрашены несколько светлее, т.к. содержат в генотипах три доминантных аллеля и один рецессивный аллель, шесть – еще светлее с двумя доминантными и двумя рецессивными аллелями в генотипах, четыре – еще более светлые, т.к. имеют лишь один доминантный и три рецессивных гена, то есть истинное расщепление будет 1:4:6:4:1. 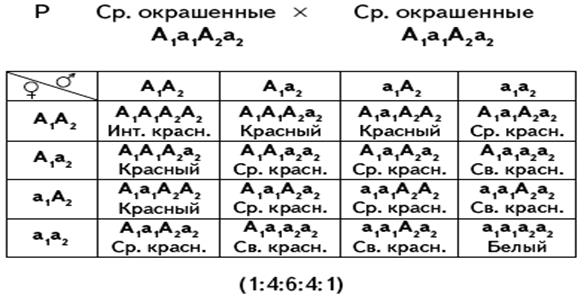
По данному варианту полимерии наследуются окраска кожи, рост и масса у человека. Подобный же механизм наследования характерен для многих количественных, в том числе и хозяйственно-ценных признаков: содержание сахара в корнеплодах свеклы, содержание витаминов в плодах и овощах, длина колоса злаков, длина початка кукурузы, плодовитость животных, молочность скота, яйценоскость кур и др.
• Полимерные гены не усиливают друг друга. В этом случае расщепление в F2 будет 15:1. Так, плоды пастушьей сумки могут быть треугольными (доминантный признак) и овальными (рецессив). Признак контролируется двумя парами полимерных генов. Если в генотипе растения имеется хотя бы один доминантный аллель из первой или второй пары полимерных генов, то форма плода у него будет треугольной (А1А2; А1а2;а1А2). Овальную форму плодов будут иметь лишь те растения, у которых в генотипе нет ни одного доминантного аллеля – а1а1а2а2.
Таким образом, накопление определенных аллелей в генотипе может привести к изменению выраженности признаков.
Билет 9
Хромосомные перестройки ( аберрации)
Хромосомные аберрации (хромосомные мутации, хромосомные перестройки) — изменения структуры хромосом. Классифицируют делеции (удаление участка хромосомы), инверсии (изменение порядка генов участка хромосомы на обратный), дупликации (повторение участка хромосомы), транслокации (перенос участка хромосомы на другую). Хромосомные перестройки носят, как правило, патологический характер и нередко приводят к гибели организма. Показано значение хромосомных перестроек в видообразовании и эволюции.
Причины хромосомных аберраций
Основной предпосылкой для возникновения хромосомных перестроек является появление в клетке двунитевых разрывов ДНК, то есть разрывов обеих нитей спирали ДНК в пределах нескольких пар оснований. Двунитевые разрывы ДНК возникают в клетке спонтанно или под действием различных мутагенных факторов: физической (ионизирующее излучение), химической или биологической (транспозоны, вирусы) природы. Двунитевые разрывы ДНК возникают запрограммированно во время профазы I мейоза, а также при созревании Т- и B-лимфоцитов во время специфической соматической V(D)J рекомбинации. Нарушения и ошибки процесса воссоединения двунитевых разрывов ДНК приводят к появлению хромосомных перестроек[3].
Делеции
Различают терминальные (утрата концевого участка хромосомы) и интеркалярные (утрата участка на внутреннем участке хромосомы) делеции. Если после образования делеции хромосома сохранила центромеру, она аналогично другим хромосомам передается при делении, участки же без центромеры как правило утрачиваются. При конъюгациигомологов во время кроссинговера у нормальной хромосомы на месте делеции в мутировавшей хромосоме образуется т. н. делеционная петля, которая компенсирует отсутствие делетированного участка.
Исследованные делеции редко захватывает протяженные участки хромосом, обычно такие аберрации летальны. Самым хорошо изученным заболеванием, обусловленным делецией, является синдром кошачьего крика, описанный в 1963 году Джеромом Леженом. В его основе лежит делеция небольшого участка короткого плеча 5 хромосомы. Для больных характерен ряд отклонений от нормы: нарушение функций сердечно-сосудистой, пищеварительной систем, недоразвитие гортани (с характерным криком, напоминающим кошачье мяуканье), общее отставание развития,умственная отсталость, лунообразное лицо с широко расставленными глазами. Синдром встречается у 1 новорожденного из 50000.
Другой интересной делецией является делеция в гене, кодирующем рецептор CCR5. Этот рецептор используетсявирусом иммунодефицита человека (ВИЧ) для распознавания своей цели — Т-лимфоцитов. Продукта гена с делецией получил название CCR5-Δ32, этот вариант CCR5 не узнается ВИЧ, и носители такой мутации к ВИЧ невосприимчивы (это порядка 10 % европейцев).
Дупликации
Дупликации появляются в результате неравного кроссинговера (в этом случае второй гомолог несет делецию) или в результате ошибки в ходе репликации. При конъюгации хромосомы с дупликацией и нормальной хромосомы как и при делеции формируется компенсационная петля.
Практически у всех организмов в норме наблюдается множественность генов, кодирующих рРНК (рибосомальную РНК). Это явление назвали избыточностью генов. Так у E. coli на рДНК (ДНК, кодирующее рРНК) приходится 0,4 % всего генома, что соответствует 5-10 копиям рибасомальных генов.
Другой пример дупликации — мутация Bar у Drosophila, обнаруженная в 20-х годах XX века Т. Морганом и А. Стертевантом. Мутация обусловлена дупликацией локуса 57.0 X-хромосомы. У нормальных самок (B+/B+) глаз имеет 800 фасеток, у гетерозиготных самок (B+/B) глаз имеет 350 фасеток, у гомозигот по мутации (B/B) — всего 70 фасеток. Обнаружены также самки с трижды повторенным геном — double Bar (BD/B+).
В 1970 году Сусумо Оно в монографии «Эволюция путем дупликации генов» разработал гипотезу об эволюционной роли дупликаций, поставляющих новые гены, не затрагивая при этом функций исходных генов. В пользу этой идеи говорит близость ряда генов по нуклеотидному составу, кодирующих разные продукты. Это трипсин и хемотрипсин,гемоглобин и миоглобин и ряд других белков.
Инверсии
Различают парацентрические (инвертированный фрагмент лежит по одну сторону от центромеры) и перицентрические (инвертированный фрагмент лежит по разные стороны от центромеры) инверсии. При инверсиях не происходит потери генетического материала, потому как таковые инверсии как правило не влияют на фенотип, но если в инверсионной гетерозиготе (то есть организме, несущем как нормальную хромосому, так и хромосому с инверсией) происходиткроссинговер, то существует вероятность формирования аномальных хроматид. В случае парацентрической инверсии образуется одна нормальная и одна инвертированная (фенотипически нормальная) хроматиды, дицентрическая хроматида с дупликацией и делецией (при расхождении хроматид она обычно разрывается на две) и ацентрическая хроматида с дупликацией и делецией (обычно утрачивается). В случае перицентрической инверсии образуется одна нормальная и одна инвертированная хроматиды, а также две хроматиды с дупликацией и делецией. Гаметы, несущие дефектные хромосомы, обычно не развиваются или погибают на ранних этапах эмбриогенеза. Но гаметы с инвертированной хромосомой развиваются в организмы, 50 % гамет которых нежизнеспособны. Т.о. мутация сохраняется в популяции.
У человека наиболее распространенной является инверсия в 9 хромосоме, не вредящая носителю, хотя существуют данные, что у женщин с этой мутацией существует 30 % вероятность выкидыша.
Транслокации
Помимо переносов участков с одной негомологичной хромосомы на другую, классифицируют также реципрокные транслокации (когда две негомологичные хромосомы обмениваются участками), Робертсоновские транслокации (при этом две негомологичные хромосомы объединяются в одну), а также транспозиции (перенос участка хромосомы на другое место на той же хромосоме).
Транслокация, реципрокная транслокация и транспозиция, которые не сопровождаются утратой генетического материала (сбалансированные транслокации), часто не проявляются фенотипически. Однако, как и в случае с инверсиями, в процессе гаметогенеза часть сформированных гамет несет летальные перестройки. Например, в случае реципрокной транслокации обычно выживает не более 50 % зигот.
Примером транслокации может служить "семейный" синдром Дауна. При этом заболевании у одного из родителей обнаруживается фенотипически не проявляющаяся транслокация 21-ой хромосомы на 14-ую. У такого человека с вероятностью в 1/4 образуются гаметы с двумя 21 хромосомами (одна свободная и одна траслоцированная). При слиянии такой гаметы с нормальной образуется трисомик по 21 хромосоме.
Другой пример — транслокация типа "Филадельфийская транслокация" между девятой и двадцать второй хромосомами. В 95 % случаев именно эта мутация является причиной одной из форм хронической лейкемии (chronic myelogenous leukemia).
Робертсоновские транслокации, возможно, являются причиной различий между числом хромосом у близкородственных видов. Показано, что два плеча 2-й хромосомы человека соответствуют 12 и 13 хромосомамшимпанзе. Возможно, 2-я хромосома образовалась в результате робертсоновской транслокации двух хромосомобезьяноподобного предка человека. Таким же образом объясняют тот факт, что различные виды дрозофилы имеют от 3 до 6 хромосом.
Робертсоновские транслокации привели к появлению в Европе нескольких видов-двойников (хромосомные расы) у мышей группы видов Mus musculus, которые, как правило, географически изолированы друг от друга. Набор и, как правило. экспрессия генов при робертсоновских транслокациях не изменяются, поэтому виды практически неотличимы внешне. Однако они имеют разные кариотипы, а плодовитость при межвидовых скрещиваниях резко понижена.
|
из
5.00
|
Обсуждение в статье: Полимерноре взаимодействие генов |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы