 |
Главная |


Типы питания прокариот
|
из
5.00
|
Прокариоты характеризуются многообразием типов питания в отличие от растений (автотрофы) и животных (ге- теротрофы). Для характеристики типов питания используют три критерия: источник углерода, источник энергии, донор электронов (водорода). По источнику углерода делят на ав- тотрофы и гетеротрофы, по источнику энергии – на фото- трофы (используют солнечный свет) и хемотрофы (получа- ют энергию за счет окислительно-восстановительных реак- ций), по донору – электроны (водорода) на литотрофы (неорганические соединения Н2, NH3, H2S, Fe2 + , CO и др.) и органотрофы (используют в качестве донора электрона ор- ганические соединения). Таким образом, выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы, хемолитоавтотрофы, хемоорганоге- теротрофы.
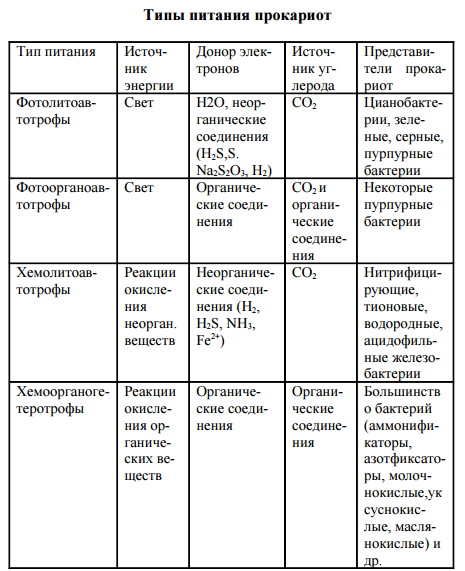
Фотолитоавтотрофы. Бактериальный фотосинтез (фоторедукция). В 1931 г. К. ван Ниль впервые доказал способность бактерий к фотосинтезу. В основе бактериального фотосинтеза лежит превращение световой энергии, поглощаемой фотосинтетическим пигментом, в биохимическую энергию макроэргических связей (АТФ) и далее использо- вание этой энергии для восстановления углекислого газа в процессе биосинтеза. У этих организмов есть пигмент бак- териохлорофилл. В клетках всех фотосинтезирующих бактерий содержатся фотосинтетические пигменты. К ним относятся особые хлорофиллы, получившие название бактериохлорофиллов а, в, с, d, и каротиноиды. По строению бактерио- хлорофиллы близки к хлорофиллу а растений. Так, бакте- риохлорофилл а отличается от хлорофилла а растений тем, что в первом его пирольном кольце в положении 2 стоит ацетильная группа СН3-СО- вместо обычной винильной группы СН2=СН-, а второе пирольное кольцо восстановле- но и содержит на два атома водорода больше. Содержание в клетке фотосинтезирующих прокариот четырех видоизменений бактериохлорофилла обусловли- вает более широкую полосу спектра поглощения световой энергии по сравнению с хлорофиллом растений. Предел спектра поглощения хлорофилла растений лежит в области 700-780 нм, в то время как фотосинтезирующие бактерии используют световую энергию длинноволновой части спектра до 1100 нм. Разница в спектрах поглощения хлоро- филлов растений и бактериохлорофиллов фотосинтезиру- ющих бактерий расширяет экологические ниши последних и позволяет фотосинтезирующим бактериям развиваться в водоемах под слоем водорослей. Помимо бактериохлорофиллов, в клетках фотосинтезирующих бактерий открыты более 20 дополнительных каро- тиноидных пигментов. Особенно разнообразны каротинои- ды в клетках пурпурных бактерий. Каротиноиды фотосин- тезирующих бактерий поглощают световую энергию ко- ротковолновой видимой области спектра с длиной волны 400-550 нм и передают эту энергию на бактериохлоро- филл. В клетке прокариот фотосинтетические пигменты нахо- дятся на инвагинациях ЦПМ – хроматофорах, визикулах (пузырьках), трубочках, тиллакоидах, ламеллах. Ван Ниль назвал этот процесс бактериальным фотосин- тезом, или фоторедукцией.
Для восстановления одной молекулы углекислого газа требуется 1 молекула АТФ (у растений 4 АТФ). В том и другом случае идет восстановление углекислого газа до углеводов. В Определителе Берджи (1974) фотосинтезирующие бактерии представлены тремя семействами (см. выше).
Фотоорганоавтотрофы Фотоорганоавтотрофы представлены немногочисленным семейством Rhodospirillaceae, включающим три рода: Rhodospirillum, Rhodopseudomonas и Rhodomicrobium. Бактерии-фотоорганоавтотрофы способны перестраивать свой обмен и одинаково успешно развиваться как на свету, так и в темноте, переходя соответственно от ана- эробного образа жизни к аэробному. На свету они ведут себя как фотоорганоавтотрофы: усваивая углекислый газ, они восстанавливают его в процессе фотосинтеза до углевода. В качестве доноров электронов несерные пурпурные бактерии используют различные органические вещества - сахара, спирты, органические кислоты, аминокислоты. Характер использования органических веществ различными фотосинтезирующими бактериями существенно раз- личается. Чаще всего органическое вещество выполняет единственную функцию донора электронов при фотоасси- миляции углекислого газа. Однако в некоторых случаях органическое вещество используется фотосинтезирующими бактериями не только в качестве донора электронов, но одновременно и как источник углерода. Попадая в темноту, пурпурные бактерии переходят к хемоорганогетеротрофному типу питания. При этом энергию для процессов жизнедеятельности они получают за счет реакций окисления органического субстрата по циклу Кребса. Непосредственным источником углерода и донором электронов для них являются органические соедине- ния субстрата. В клетках бактерий этой группы имеется универсальный набор дыхательных ферментов (НАД- и ФАД-дегидрогеназы и цитохромы), обеспечивающих им возможность перехода от анаэробного образа жизни на свету к аэробному в темноте и, соответственно, от авто трофного типа питания к гетеротрофному. Таким образом, среди разных групп прокариот нет, да и не может быть, резкой границы между различными типами питания.
ХемолитоавтотрофыХемолитоавтотрофы представлены микроорганизмами, способными в качестве основного источника углерода усваивать углекислый газ и синтезировать в клетке органические соединения, используя энергию реакций окисления неорганического субстрата. Для хемолитоавтотрофов неорганические вещества субстрата выступают донорами электронов в реакциях энергетического метаболизма и в процессе хемоассимиляции углекислого газа. Заслуга открытия процесса хемосинтеза принадлежит С. Н. Виноградскому. Он определил химизм как жизнь без органики. Им впервые была показана возможность образования органических веществ из неорганических в клетках бактерий, помимо процесса фотосинтеза. К хемолитоавтотрофам относится большинство видов нитрифицирующих, тионовых бактерий, некоторые виды из группы одноклеточных железобактерий и водородные бактерии. Хемолитоавтотрофные бактерии характеризуются специфичностью в отношении использования окисляемого субстрата. Нитрифицирующие бактерии для процесса хемоассимиляции углекислого газа получают энергию от окисления аммиака и нитритов. Процесс нитрификации проходит в две фазы. Первая фаза заключается в аэробном окислении аммиака до нитритов нитрозными бактериями родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira: NН3 + 11 /202 = HN02 + Н2О + 274,7 кДж. Вторая фаза нитрификации предусматривает окисление нитритов в нитраты нитратными бактериями родов Nitrobacter, Nitrospina, Nitrococcus: HN02 + 11 /202 =НNО3 + 87,5 кДж. При процессах окисления неорганического субстрата выделяется сравнительно небольшое количество энергии, и усваивается она клеткой с низким КПД, всего 5-10%. Поэтому для получения энергии на процессы жизнедеятельности нитрифицирующим бактериям приходится перерабатывать огромное количество субстрата. К хемолитоав- тотрофам относится большинство видов тионовых бактерий: Thiobacillus denitrificans, Т. thiooxidans, Т. acidophilus, Thiomicrospira pelophila и др. Они ведут процесс хемоасси- миляции углекислого газа, получая энергию за счет окисления восстановленных или частично восстановленных соединений серы, сероводорода, элементарной серы, тиосульфата и сульфита. Конечным продуктом окисления обычно является сульфат.
Некоторые виды тионовых бактерий (Thiobacillus ferrooxidans) способны получать энергию за счет окисления не только соединений серы, но и закисного железа (Fe2+) (разрушение труб). Хемолитоавтотрофный тип питания характерен для некоторых одноклеточных ацидофильных железобактерий – Leptospirillum ferrooxidans, Thiobacillus ferrooxidans и представителей рода Sulfolobus. Оптимальный рост ацидофильных железобактерий наблюдается при рН ниже 4,5 (2-3). Они используют энергию окисления Fe2+ до Fе3+ для ассимиляции С02, который служит основным или единственным источником углерода. Реакции окисления железа сопровождаются незначительным выделением энергии, поэтому железобактерии перерабатывают большие количества субстрата: 2Fe2+ + 1/202 + 2Н+ = 2FеЗ+ + Н2О + 33 кДж. Хемолитоавтотрофные бактерии в природе являются геологическими агентами. Они принимают участие в процессах образования полезных ископаемых и осуществляют важнейшие звенья круговорота азота, серы, железа. К факультативным хемолитоавтотрофам относятся водородные бактерии, являющиеся представителями 20 различных родов – Рsеudоmоnаs, Аlcаligеnеs, Nocardia и др. Они способны осуществлять ферментативное окисление водорода кислородом воздуха с образованием воды. Водородные бактерии ферментом дегидрогеназой активируют молекулярный водород и далее используют его для получения энергии и в качестве донора электронов для восстановления углекислого газа до углевода: 6Н2 + 202 + С02 = (СН2О) + 5Н2О. Являясь факультативными хемолитоавтотрофами, водородные бактерии в качестве источника энергии и углеро- да способны использовать и различные органические со- единения (сахара, органические кислоты, спирты). Таким образом, водородные бактерии могут служить еще одним 87 примером возможного перехода микробной клетки от одного типа питания к другому – от хемолитоавтотрофного к хемоорганогетеротрофному. Бактерии, переходящие от одного типа питания к другому, получили название миксо- трофов.
Хемоорганогетеротрофы. К хемоорганогетеротрофам относится большинство прокариот. Источником углерода для них являются самые разнообразные органические соединения. Энергию для жизнедеятельности они получают за счет окислительно- восстановительных реакций органического субстрата, и донором электронов в реакциях метаболизма также выступают различные органические вещества. Хемоорганогетеротрофы наиболее широко распространены в природе. Им принадлежит роль санитаров нашей планеты, так как они ведут процессы минерализации самых разнообразных, подчас сложных органических веществ. Помимо органических соединений как источника углерода, хемоорганогетеротрофы нуждаются в углекислом газе для реакций карбоксилирования промежуточного обмена. Хемоорганогетеротрофные микроорганизмы подразделяют на сапрофитов и паразитов. Сапрофиты потребляют органические вещества опада. Паразиты живут за счет органических веществ живой клетки. Выделяют факультативных и облигатных паразитов. Факультативные паразиты развиваются на обычных органических средах, но, попадая в клетку-хозяина, переходят к паразитическому образу жизни. К ним относится большинство патогенных бактерий, вызывающих заболевания человека, – возбудители пневмонии, менингита, гонореи, дизентерии, брюшного тифа, сибирской язвы, коклюша, туберкулеза и др. Облигатные (строгие) паразиты развиваются исключительно за 88 счет органических веществ клетки-хозяина. Типичным примером облигатных паразитов являются риккетсии и вирусы. В основе всех вышерассмотренных типов питания прокариот лежат различные сочетания их энергетического и конструктивного метаболизма. Всем типам питания соот- ветствуют определенные группы прокариотных организ- мов. Основной массе бактерий присущ один вполне определенный (облигатный) тип питания. Однако среди многообразного мира прокариот встречается много видов бактерий, способных переходить от одного типа питания к другому. Так, среди цианобактерий, зеленых и пурпурных бактерий имеются виды, переходящие от фотолитоавто- трофного типа питания к фотоорганогетеротрофному. Некоторые виды обширного рода Thiobacillus способны переходить от хемолитоавтотрофного типа питания к хемоорга- ногетеротрофному. Эта особенность питания прокариот обусловливает широкие возможности существования микроорганизмов в различных условиях среды и значительно расширяет их экологические ниши.
|
из
5.00
|
Обсуждение в статье: Типы питания прокариот |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы