 |
Главная |


Миелиновые нервные волокна
|
из
5.00
|
Миелиновые нервные волокна значительно толще безмиелиновых. Диаметр поперечного сечения их колеблется от 1 до 20 мк. Они также состоят из осевого цилиндра, одетого оболочкой из леммоцитов, но диаметр осевых цилиндров этого типа волокон значительно больше, а оболочка сложнее. В сформированном миелиновом волокне принято различать два слоя оболочки: внутренний, более толстый, - миелиновый слой (рис.10), и наружный, тонкий, состоящий из цитоплазмы леммоцитов и их ядер.
Миелиновый слой содержит в своем составе липоиды, а поэтому при обработке волокна осмиевой кислотой он интенсивно закрашивается в темно-коричневый цвет. Все волокно в этом случае представляется однородным цилиндром, в котором на определенном расстоянии друг от друга располагаются косо ориентированные светлые линии - насечки миелина (incision myelini), ил и насечки Шмидта-Лантермана. Через некоторые интервалы (от нескольких сотен микронов до нескольких миллиметров) волокно резко истончается, образуя сужения - узловые перехваты, или перехваты Ранвье. Перехваты соответствуют границе смежных леммоцитов. Отрезок волокна, заключенный между смежными перехватами, называется межузловым сегментом, а его оболочка представлена одной глиальной клеткой.
В процессе развития миелинового волокна осевой цилиндр, погружаясь в леммоцит, прогибает его оболочку, образуя глубокую складку.

Рис. 10. Схема нейрона. 1 – тело нервной клетки; 2 – осевой цилиндр; 3 – глиальная оболочка; 4 – ядро леммоцита; 5 – миелиновый слой; 6 – насечка; 7 – перехват Ранвье; 8 – нервное волокно, лишенное миелинового слоя: 9 – двигательное окончание; 10 – миелиновые нервные волокна, обработанные осмиевой кислотой.
По мере погружения осевого цилиндра оболочка леммоцита в области щели сближается и ее два листка соединяются друг с другом своей внешней поверхностью, образуя двойную мембрану - мезаксон (рис.11).
При дальнейшем развитии миелинового волокна мезаксон удлиняется и концентрически наслаивается на осевой цилиндр, вытесняя цитоплазму леммоцита и образуя вокруг осевого цилиндра плотную слоистую зону - миелиновый слой (рис.12). Так как оболочка леммоцита состоит из липидов и белков, а мезаксон представляет собой ее двойной листок, то естественно, что миелиновая оболочка, образованная его завитками, интенсивно окрашивается осмиевой кислотой. В соответствии с этим под электронным микроскопом каждый завиток мезаксона виден как слоистая структура, построенная из белков и липидов, расположение которых типично для мембранных структур клеток. Светлый слой имеет ширину около 80-120 Ǻ и соответствует липоидным слоям двух листков мезаксона. Посредине и по поверхности его видны тонкие темные линии, образованные молекулами белка.
Рис. 11. Схема развития миелинового волокна. 1 – контакт аксолеммы и оболочки леммоцита; 2 – щель; 3 – аксолемма и оболочка леммоцига; 4 – цитоплазма леммоцита; 5 – мезаксон
Шванновской оболочкой называется периферическая зона волокна, содержащая оттесненную сюда цитоплазму леммоцитов (шванновских клеток) и их ядра. Эта зона при обработке волокна осмиевой кислотой остается светлой. В области насечек между завитками мезаксона имеются значительные прослойки цитоплазмы, благодаря чему клеточные мембраны располагаются на некотором расстоянии друг от друга. Больше того, как видно на рис.188, листки мезаксона в этой области также лежат неплотно. В связи с этим при осмировании волокна эти участки не окрашиваются.
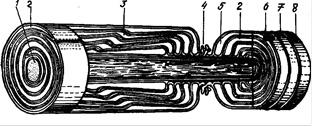
Рис. 12. Схема субмикроскопического строения миелинового нервного волокна: 1 – аксон; 2 – мезаксон; 3 – насечка миелина; 4 – узел нервного волокна; 5 – цитоплазма нейролеммоцита; 6 – ядро нейролеммоцита; 7 – нейролемма; 8 – эндоневрий
На продольном сечении вблизи перехвата видна область, в которой завитки мезаксона последовательно контактируют с осевым цилиндром. Место прикрепления самых глубоких завитков его наиболее удалено от перехвата, а все последующие завитки закономерно расположены ближе к нем у (см. рис.12). Это легко понять, если представить себе, что закручивание мезаксона идет в процессе роста осевого цилиндра и одевающих его леммоцитов. Естественно, что первые завитки мезаксона оказываются короче, чем последние. Края двух смежных леммоцитов в области перехвата образуют пальцеобразные отростки, диаметр которых равен 500 Ǻ. Длина отростков различна. Переплетаясь между собой, они образуют вокруг осевого цилиндра своеобразный воротничок и попадают на срезах то в поперечном, то в продольном направлении. В толстых волокнах, у которых область перехвата относительно коротка, толщина воротничка из отростков шванновских клеток больше, чем в тонких волокнах. Очевидно, аксон тонких волокон в перехвате более доступен для внешних воздействий. Снаружи миелиновое нервное волокно покрыто базальной мембраной, связанной с плотными тяжами коллагеновых фибрилл, ориентированных продольно и не прерывающихся в перехвате - невралеммой.
Функциональное значение оболочек миелинового нервного волокна в проведении нервного импульса в настоящее время недостаточно изучено.
Осевой цилиндр нервных волокон состоит из нейроплазмы - бесструктурной цитоплазмы нервной клетки, содержащей продольно ориентированные нейрофиламенты и нейротубулы. В нейроплазме осевого цилиндра лежат митохондрии, которых больше в непосредственной близости к перехватам и особенно много в концевых аппаратах волокна.
С поверхности осевой цилиндр покрыт мембраной - аксолеммой, обеспечивающей проведение нервного импульса. Сущность этого процесса сводится к быстрому перемещению локальной деполяризации мембраны осевого цилиндра по длине волокна. Последнее определяется проникновением в осевой цилиндр ионов натрия (Nа+), что меняет знак заряда внутренней поверхности мембраны на положительный. Это, в свою очередь, повышает проходимость ионов натрия в смежном участке и выход ионов калия (К+) на внешнюю поверхность мембраны в деполяризованном участке, в котором восстанавливается при этом исходный уровень разности потенциалов. Скорость движения волны деполяризации поверхностной мембраны осевого цилиндра определяет быстроту передачи нервного импульса. Известно, что волокна с толстым осевым цилиндром проводят раздражение быстрее тонких волокон. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1-2 м/сек, тогда как толстые миелиновые - 5-120 м/сек.
Нервные окончания
Конечные разветвления нейронов образуют нервные окончания, которые контактируют с тканями (органами), воспринимающими нервные импульсы из центра, либо с тканями (органами), передающими воздействие внутренней и внешней среды и способствующими образованию нервных импульсов, направленных в центр. В связи с этим различают эффекторы и рецепторы.
Эффекторы, или двигательные и секреторные нервные окончания, проводят нервные импульсы от мозга к рабочему органу (мышца, железа) и регулируют его функцию, а рецепторы, или чувствительные нервные окончания, передают нервные импульсы, вызванные различными раздражителями извне (кожа, скелетная мускулатура, внутренние органы).
Эффекторы - двигательные и секреторные нервные окончания - имеют различное строение. На миоцитах неисчерченной мышечной ткани (сарколемма) они представлены в виде небольших концевых пуговок, из сплетения нейрофибрилл в виде петли, в центре которой находится скопление аксоплазмы. Субмикроскопически это окончание имеет структуру синапса (рис. 13) с типичными особенностями (см. ниже). Очень характерно, что одно нервное волокно, подходя к миоциту, делится на ветви, образующие подобие сплетения и оканчивающиеся концевыми пуговками на сарколемме. Этим увеличивается контакт нейрона с миоцитом. В месте контакта, как уже доказано, нервное окончание вырабатывает особые продукты обмена, называемые медиаторами (холин - и адренергические вещества), которые способствуют передаче импульса на рабочий орган (ткань, клетка), а также и сами могут быть причиной возбуждения и специфической работы.

Рис. 13. Схема субмикроскопического строения двигательного нервного окончания на миоците: 1 – постсиналтнческая мембрана миоцита; 2 – митохондрия; 3 – светлые пресинаптические пузырьки; 4 – пресинаптическая мембрана нейрита
Это проверяется введением их в кровь, лимфу, когда медиаторы проявляют свое действие на другие органы (ткани).
В связи с отмеченным, полагают, что увеличение степени разветвления одного нервного волокна для образования контакта с одной мышечной клеткой способствует выработке большого количества медиатора и лучшей передаче нервного импульса.
Методом перерезки при последующей дегенерации нервных волокон, о чем будет сказано позже, доказано, что неисчерченная мускулатура иннервируется вегетативной нервной системой (постганглионарные нервные волокна).
На сердечных миоцитах (мышечные клетки миокарда сердца) в основном наблюдаются такие же нервные окончания, как и на миоцитах неисчерченной мышечной ткани. Источником иннервации также является вегетативная нервная система, причем трудно отличить окончания постганглионарных симпатических и парасимпатических волокон.
Окончания на неисчерченной и сердечной исчерченной мышечной ткани являются экстрацеллюлярными (внеклеточными), а по утверждению Ж. Буке (1913), они проникают в саркоплазму миоцитов.
Скелетная мускулатура получает иннервацию от мозга, который посылает нервные волокна к мышечным волокнам.

Рис. 14. Схема субмикроскопического строения нейромышечного окончания: 1 – цитоплазма нейролеммоцита; 2 – ядро нейролеммоцита; 3 – нейролемма, переходящая в наружный слой сарколеммы; 4 – аксон; 5 – сарколемма; 6 – концевые веточки нервного волокна в продольном и поперечном сечениях; 7 – митохондрии в аскоплазме: 8 – первичное синаптическое пространство; 9 – саркосомы; 10 – вторичное синаптическое пространство; 11 – светлые пресинаптические пузырьки; 12 – пресинаптическая мембрана; 13 – постсинаптическая мембрана; 14 – ядро мышечного волокна; 15 – миофибрилла, состоящая из миофиламентов
Как показывает световой микроскоп, в месте контакта с мышечным волокном нервное волокно теряет миелиновую оболочку, эндоневрий сливается с сарколеммой, которая расщепляется на два листка и включает разветвление аксона в виде петелек и пластинок из нейрофибрилл и ядер нейролеммоцитов. Окончания на мышечных волокнах скелетной мускулатуры называются нейромышечными (концевые моторные - бляшками), которые заложены внутрь сарколеммы и изолированы своим основанием - подошвой - от саркоплазмы (М.Д. Лавдовский, 1884-1885; К.А. Лавров, 1941). Ноэль (1932) доказал, что митохондрии подошвы концевой моторной бляшки близки по структуре к наблюдаемой в нейролеммоцитах и резко разнятся от митохондрий исчерченного мышечного волокна. Кроме того, при некоторых патологических процессах отмечались (Л.И. Фалин, 1954; Е.К. Плечкова, 1955) реактивные изменения в ядрах нейролеммоцитов (размножение, вакуолизация) подошвы концевых моторных бляшек, а мышечные волокна оставались без видимых изменений.
На электронограммах также можно наблюдать контакт нервных и мышечных волокон в области концевых моторных бляшек. Субмикроскопически каждая из них (рис.14) состоит из нервного волокна, нейролемма которого сливается с сарколеммой, аксон образует нервный полюс, т.е. концевую веточку (пальцеообразные выросты) и вместе с пограничной мембраной мышечного волокна погружается в саркоплазму. Аксолемма окружает разветвления аксона и образует пресинаптическую мембрану с пальцеобразными выростами. Между аксоплазмой и примыкающей к ней пресинаптической мембраной располагается первичное синаптическое пространство, а между пальцеобразными складками той же мембраны - вторичное синаптическое пространство. Эти щелевидные пространства заполнены гомогенным веществом. Кнаружи от пресинаптической мембраны залегает цитолемма мышечного волокна, или постсинаптическая мембрана. В аксоплазме разветвлений аксона содержатся пресинаптические пузырьки и митохондрии, которые в отличие от наблюдаемых в исчерченном мышечном волокне имеют не поперечные, а продольные митохондриальные гребни.
Установлено, что в области концевой моторной бляшки (пресинаптической части) вырабатывается ацетилхолин, а в области ее подошвы (постсинаптической части) - ацетилхолинэстераза, способствующая гидролизу ацетилхолина (В.В. Португалов, 1955). Рядом экспериментов и наблюдений доказано, что концевые моторные бляшки обладают очень высокой реактивностью.
Эффекторы наблюдаются на клетках эпителия (нервно-эпителиальные окончания) и мезенхимы. На клетках реснитчатого эпителия заканчиваются постганглионарые нервные волокна симпатических звездчатых узлов (Лашков, 1948). Нервные волокна проникают в эпителий, оплетают каждую его клетку и заканчиваются на ней концевой пуговкой (пуговчатое утолщение) в месте расположения запирающей зоны (замыкающие пластинки). Подобные окончания образуются и на клетках железистого эпителия (К.А. Арнштейн, 1895; Н.И. Зазыбин, 1933). Это нервно-железистые окончания. Пуговчатыми окончаниями симпатических нервных волокон обладают также остеобласты, причем при их дифференцировке в остеоциты нервное окончание исчезает. На липоцитах и на элементах мезенхимы также установлены нервные окончания.
Все окончания на элементах соединительной ткани имеют трофическое значение.
Рецепторы (чувствительные нервные окончания) встречаются в двух основных формах: свободные и связанные с дополнительными образованиями (клетками).
Свободные нервные окончания образуются миелиновыми нервными волокнами, которые теряют все оболочки при вхождении в иннервируемую ткань. Они состоят из аксонов, переходящих в разветвления, заканчивающиеся концевыми колечками и пластинками, образованными нейрофибриллами и аксоплазмой. По предположению И.Н. Филимонова (1955), каждое разветвление одето цитоплазмой нейролеммоцитов. Отсюда понятие “свободные нервные окончания" является относительным. Эта форма нервных окончаний наблюдается в эпидермисе, во внешнем влагалище волоса, на волокнах неисчерченной и сердечной исчерченной мускулатуры и т.д.
В эпидермисе, например, свободные окончания образованы миелиновыми волокнами, которые подходят вплотную к базальной мембране, теряют миелин, разветвляются и заканчиваются концевыми пуговками и пластинками на клетках, причем эти разветвления не выходят за пределы блестящего слоя. В самых поверхностных слоях эпидермиса нервные нити приобретают варикозные расширения (вздутия) и фрагментируются, что свидетельствует о постоянном раздражении, воспринимаемом конечными разветвлениями описываемых волокон. Окончания в эпидермисе относятся к экстероцепторам (воспринимают болевые раздражения).
В наружном корневом влагалище волоса конечные разветвления аксона приобретают вид частокола, окружающего волос, т.е. параллельного его оси. Это окончания волосяных фолликулов.
Ко второй группе рецепторов относятся такие нервные образования, аксоны которых связаны со специальными осязательными (чувствительными) клетками, по-видимому, возникающими из нейролеммоцитов.
У всех млекопитающих в эпидермисе обнаружены осязательные мениски, которые впервые были найдены в пятачке свиньи (рис.15). Они состоят из светлых осязательных клеток овоидной или овоидно-эллиптической формы, которые лежат на нервных пластинках типа розеток. Осязательные мениски - типичные экстероцепторы (проводят тактильную чувствительность).
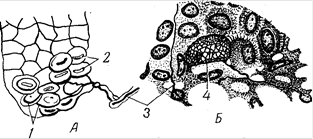
Рис. 15. Рецепторные нервные окончания в эпителии: А – кожи свиньи; Б – из эймерова органа крота; 1 – осязательные клетки; 2 – осязательные мениски; 3 – нервные волокна; 4 – сетчатая структура вокруг осязательной клетки
К описываемым рецепторам относятся и осязательные тельца продолговато эллиптической формы (рис.16). Они лежат в сосочковом слое кожи под эпидермисом перпендикулярно его поверхности и состоят из тонкой соединительной капсулы осязательного тельца. К ней подходит миелиновое нервное волокно, оно, как правило, под капсулой теряет миелиновую оболочку, разветвляется и заканчивается пластинками из нейрофибрилл, на которых лежат олигодендроглиоциты - чувствительные клетки. Осязательные тельца находят и в слизистых оболочках, например в надгортаннике.
Другой разновидностью второй группы рецепторных окончаний являются капсулированные нервные тельца. Они имеют сильно развитую капсулу и обладают разнообразным строением. До последнего времени сюда же относили и осязательные тельца.
Наиболее типичные капсулированные нервные тельца - это концевые нервные тельца, генитальные, пластинчатые тельца и др.

Рис. 16. Осязательные тельца: А – из сосочка кожи пальца человека; Б – из кожи пальца человека; 1 – эпидермис; 2 – основное волокно; 3 – добавочное волокно; 4 – расширение основного волокна; 5 – олигодендроглиоциты (осязательные клетки); 6 – веточка нервного волокна, выходящая из тельца; 7 – пластинка
Концевые нервные тельца (слизистые оболочки рта, языка, мочевыводящих путей) состоят из довольно толстой соединительнотканной капсулы, к которой подходит миелиновое нервное волокно, теряющее под капсулой миелин и в виде ленты или пластинки располагающееся в центре тельца. Нередко от пластинки отходят разветвления в виде шипиков. Пластинка окружается клетками типа нейролеммоцитов. В целом это так называемая внутренняя луковица.
Более сложно устроены генитальные тельца (слизистые оболочки половых органов, рта и дыхательного аппарата). Заметим, что такое название является неудачным, так как эти тельца не связаны со специфическими ощущениями от половых органов, как раньше предполагали. Это скорее аппараты тактильного чувства. Их внутренняя луковица состоит из разветвления в виде сетей и клубочков, образованных несколькими миелиновыми нервными волокнами, подходящими к тельцу.
 Широко распространены в организме пластинчатые тельца. Это почти макроскопические образования (сетчатый слой кожи, подкожная основа, брыжейка, паренхима многих желез - слюнных, поджелудочной, предстательной, надпочечника).
Широко распространены в организме пластинчатые тельца. Это почти макроскопические образования (сетчатый слой кожи, подкожная основа, брыжейка, паренхима многих желез - слюнных, поджелудочной, предстательной, надпочечника).
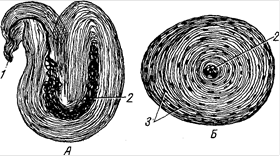
Рис. 17. Капсулированные нервные окончания: А – продольный разрез пластинчатого тельца из кожи человека; Б – поперечный срез пластинчатого тельца из кожи пальца человека; 1 – нервное волокно; 2 – внутренняя луковица; 3 – клетки пластинок капсулы
Тельца (рис. 17) состоят из очень толстой капсулы - наружной луковицы - результат наслоения нескольких десятков пластинок плотной соединительной ткани (или глии) с клетками и тканевой жидкостью между пластинками. Внутренняя луковица образована видоизмененными нейролеммоцитами и аксоном в виде длинной пластинки, иногда снабженной шипиками и выростами, а сам аксон ветвится. Кроме основного волокна, к тельцу подходит тонкое миелиновое или безмиелиновое добавочное волокно, аксон которого проникает во внутреннюю луковицу и разветвляется вокруг основного волокна. Раньше добавочные волокна относили к вегетативному отделу нервной системы, а в настоящее время (методом перерезки нервных волокон) установлена их цероброспинальная природа. Субмикроскопическое строение пластинчатых телец отличается от оптического некоторыми деталями (рис.18). По мнению большинства авторов, пластинчатые тельца реагируют на давление (проприоцепторы).
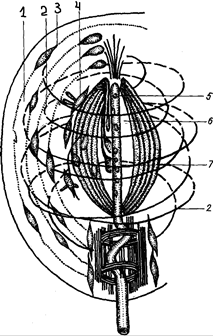
Рис. 18. Схема субмикроскопического строения пластинчатого тельца: 1 – слоистая капсула; 2 – спиральные коллагеновые волокна; 3– фиброцит; 4 – вторично чувствующие клетки с ресничками; 5 – дендрит рецепторного нейрона; 6 – внутренняя колба; 7 – синаптические контакты аксонов вторично чувствующих клеток с дендритами рецепторного нейрона
К капсулированным нервным окончаниям относят чувствительные концевые аппараты волокон скелетной мускулатуры - нервно-мышечные веретена. Строение их таково: одно или несколько очень тонких мышечных волокон заключено в специальную соединительно-тканную капсулу в форме овала или веретена. Толстые миелиновые нервные волокна прободают капсулу, теряют миелин, ветвятся в сопровождении специальных, по-видимому, глиальных клеток, подходят к мышечным волокнам и спирально их обвивают. Часть нервных нитей заканчивается спиральными пластинками на сарколемме мышечного волокна. Нервно-мышечные веретена передают в центр сигналы о состоянии тонуса мышечных волокон.
Исследованиями установлено наличие в кровеносной системе и в стенках ряда внутренних органов млекопитающих особых некапсулированных нервных окончаний - хеморецепторов, улавливающих изменения химического состава крови (общее сонное и грудное аортальное сплетения). Они состоят из разветвлений аксонов, сопровождаемых специальными клетками (глиальными).
Предполагают, что разные пороги раздражения общего сонного и грудного аортального сплетений объясняются разным количеством имеющихся в них специальных клеток.
В заключение следует отметить, что рецепторы обладают одной особенностью строения, изученной Б.И. Лаврентьевым (1941). Было установлено, что одно и то же чувствительное волокно на периферии ветвится и образует нервные окончания на различных тканевых образованиях разных органов, например в эпителии, в неисчерченных мышцах и на сосудах в желудочно-кишечном тракте, в сердечной исчерченной мускулатуре и на венечных сосудах. Такие окончания Б.И. Лаврентьев назвал “поливалентными" рецепторами. Нужно полагать, что к такому роду рецепторам относятся и те, которые способны к осуществлению аксон-рефлексов. Полагают, что аксон-рефлекс совершается через разветвления одного чувствительного нервного волокна, ветви которого связывают эпидермис с кровеносными сосудами. Раздражение эпидермиса вызывает расширение сосудов.
Положение Б.И. Лаврентьева о “поливалентных” рецепторах подтверждает правильность предположений авторов об аксон-рефлексе и определяет механизм его осуществления.
Синапсы
До последнего времени существовали две точки зрения о взаимосвязи между нейронами.
По мнению одних (Г. Гельд, 1895; К. Гольджи, 1880; С. Апати, 1897; А. Бете, 1903), использовавших материал беспозвоночных (особенно опыты Бете с крабом), нейрофибриллы - основной путь проведения нервных импульсов, а нейроцит - сгущение его цитоплазмы, имеющей трофическое значение. Было показано, что нейрофибриллы непрерывно переходят из одного нейрона в другой. Так возникла теория нейропиля (фибриллярной непрерывности, или нейрофибриллярной решетки). Применительно к высшим животным эта теория оказалась умозрительной, так как ее представителям не удалось убедить ученых в справедливости своего мнения из-за отсутствия демонстративных картин, типичных для беспозвоночных (крупные нервные клетки), что не типично для позвоночных, особенно высших (мелкие нервные клетки).
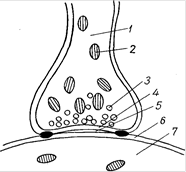
Рис.19.Схема строения синапса: 1 – пресинаптический полюс; 2 – митохондрии; 3 – светлый пресинаптический пузырек; 4 – пресинаптическая мембрана; 5 – синаптическая щель; 6 – Постсинаптическая мембрана; 7 – Постсинаптическая часть (на пре- и постсинаптической мембранах видны утолщения типа десмосом)
По мнению других (Р. Кахал, 1907; Б.И. Лаврентьев, 1939 и др.), между нейронами существует контакт - синапс (от греч. synapsis - соединение, связь). Термин дан английским физиологом Ч. Шеррингтоном. Впервые Р. Кахал установил синапсы в мозжечке: контактные связи разветвлений корзинчатых нейроцитов (вставочные нейроциты) с телами грушевидных нейроцитов (ганглиозный слой). Синапсы осуществляются передачей нервного импульса от нейрита чувствительного нейроцита к дендритам или телу корешкового (вставочного) нейроцита, а также от дендрита одного нейроцита к дендриту другого или от нейрита к нейриту двух нейроцитов. Кроме того, передача нервных импульсов может осуществляться от корешкового нейроцита к эпителиальным или мышечным клеткам.
Отсюда различают следующие синапсы: аксодендритические (контакт с дендритом последующего нейроцита), аксосоматические (контакт с телом последующего нейроцита), аксоаксональные (контакт между нейритами двух нейроцитов), дендродендритические (контакт между дендритами двух нейроцитов), аксоэпителиальные и аксомышечные (соответственно контакт корешковых нейроцитов с эпителио- и миоцитами).
Разные виды синапсов более или менее широко распространены в нервной системе. Так, аксосоматические синапсы имеют широкое распространение в нервной системе (корзинки нервных волокон на теле грушевидных нейроцитов, нейроны спинного, головного мозга, коры большого мозга, вегетативной нервной системы). Кроме того, известно, что нейрит одного нейрона, например чувствительного, может давать разветвления и оканчиваться на многих других нейронах (до 10000 синапсов). В свою очередь с одним и тем же нейроном, например корешковым нейроцитом, могут образовывать синапсы также много (1200-1800) нейронов.
Прижизненное существование синапсов у позвоночных впервые установлено в лаборатории Б.И. Лаврентьева на примере изучения внутрисердечных узлов амфибий (лягушка, жаба) с использованием метода, предложенного в 1939 г. Е.М. Граменицким (растянутая стенка предсердий на пробковом каркасе).
Прижизненными наблюдениями над синапсами и изменениями в них при тех или иных воздействиях, например ультракоротких волн, сопровождаемых возбуждением и как следствием этого желатинизацией, усилением гранулообразования (окраска янусгрюном, нейтральротом), доказано существование синапсов. Одновременно установлено совпадение наблюдаемого прижизненно и на фиксированных, а также окрашенных объектах. Субмикроскопически синапс состоит из пресинаптической, постсинаптической частей и синаптической щели. В пресинаптическую часть входит освобожденный от миелина аксон передающего нейрона, окруженный аксолеммой и содержащий в аксоплазме много митохондрий и до 3 млн. пресинаптических пузырьков диаметром 20-50 нм. В пузырьках находится медиатор, чаще всего ацетилхолин. Постсинаптическая часть представлена дендритом или телом воспринимающего нейрона, эпителиоцитом или миоцитом. Синаптическая щель расположена между пресинаптической и постсинаптической мембранами (рис. 19). Пресинаптическая мембрана - аксолемма передающего нейрона с пресинаптическим уплотнением. Элементы глии не внедряются в синаптическую щель. Постсинаптическая мембрана - цитолемма воспринимающего нейрона или другой клетки с постсинаптическим уплотнением. В ней находятся холинорецепторы (воспринимают действие ацетилхолина). Биохимическая активность в передаче нервного импульса принадлежит аксону передающего нейрона (пресинаптической части). Установлено, что перед передачей нервного импульса с нейрона на нейрон, при возбуждении передающего нейрона (пресинаптическая часть) пресинаптические пузырьки лопаются, медиатор изливается в синаптическую щель, возбуждает холинорецепторы постсинаптической мембраны и снижает ее электрический потенциал до критического. В результате этого в дендрит или тело воспринимающего нейрона (или другой клетки) усиленно проникают ионы натрия и в смежной зоне с постсинаптической мембраной возникает потенциал действия, т.е. нервный импульс. Передача нервных импульсов осуществляется всегда в одном направлении. Все это в значительной мере подтверждает справедливость взглядов представителей теории синаптических, контактных связей между нейронами. Доказано, что синаптические связи между нейронами (чувствительным, двигательным и др.) осуществляются при участии элементов нейроглии - синаптоглии (Куто, 1953). Это положение показывает, что в структуре и в функциональных проявлениях межнейронных синапсов и нервных окончаний имеются общие черты, так как и они, как правило, состоят из разветвлений аксонов, сопровождаемых нейроглией. Значение теории синапсов заключается в том, что она правильно объясняет полярность нейронов (направленность нервных импульсов от дендритов через тело к нейриту). Это положение подтверждается и данными о гистогенезе нервной ткани с установлением связи между нейронами и иннервируемыми ими тканями. Итак, нервная система едина, она состоит из нейронов и нейроглии, связанных между собой морфологически и функционально. Отсюда выдвинутая в свое время нейронная теория (В.М. Бехтерев, В. Гис, В. Вальдейер, Р. Кахал), которая трактовала, что нейрон - автономная единица нервной системы, не может быть признана вполне правильной. Рациональным в нейронной теории является то, что она подчеркивает функциональное значение нейрона в целом и в известной мере объясняет взаимоотношение центров и проводников в составе нервной системы. Накопившиеся в последующем данные позволяют внести дополнения в классическую нейронную теорию. Эти данные свидетельствуют о том, что нейрон и нейроглия взаимосвязаны, нейроны и глия очень реактивны к разнообразным внешним воздействиям и изменяются, но вместе с тем обладают значительными резервными возможностями для компенсации и восстановления нарушенных функций.
|
из
5.00
|
Обсуждение в статье: Миелиновые нервные волокна |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы