 |
Главная |


Синтез и катаболизм пуриновых оснований
|
из
5.00
|
Последовательность реакций, которая ведет к образованию пуриновых нуклеотидов, начинается с синтеза5-фосфорибозил-1-дифосфата — ФРДФ (он же пирофосфат — ФРПФ), являющегося общим предшественником фосфорибозы в синтезе пуриновых и пиримидиновых нуклеотидов (рис.8).
Перенос амидной группы глицина на ФРДФ с образованием 5-фосфорибозил-1-амина катализирует регуляторный фермент синтеза пуриновых нуклеотидовФРДФ-амидотрансфераза.
В синтезе пуриновых нуклеотидов не образуется свободное азотистое основание, а пуриновое кольцо формируется на остатке рибозо-5-фосфата при участии молекулыглицина, амидного азота глутамина, a-NH2-группы аспарагина, СО2 и одноуглеродных производных:метенил- и формил-Н4-фолата .
Синтезируется первый пуриновый нуклеотид — инозиновая кислота (IMP), которая двумя последовательными реакциями может превращаться в AMФ или в ГМФ.

Рисунок 8
Печень является основным местом образования пуриновых нуклеотидов. Она снабжает пуринами ткани, не способные к их синтезу: эритроциты, полиморфно-ядерные лейкоциты и частично мозг.
Синтез нуклеозиддифосфатов (НДФ) и нуклеозидтрифосфатов (НТФ) происходит при участии АТР и ферментов:нуклеозидмонофосфат (НМФ)- и НДФ-киназ. Например, АМР-киназа (миокиназа) катализирует реакцию:
AMP+ATФ « 2AДФ
ГМФ-киназа катализирует:ГМФ + АТР ® ГДФ +ADP
НДФ-киназа превращает НДФ в НТФ:
НДФ +ATФ ® НТФ +АДФ
AMФ, ГМФ и ИМФ ингибируют ключевые реакции своего синтеза помеханизму отрицательной обратной связи. Первые 2 фермента отвечают за скорость синтеза пуриновых нуклеотидов по основному пути, и их ингибирование происходит лишь при одновременном повышении концентрации
AMФ, ГМФ
Скорость синтеза ФРДФ зависит от 2 факторов: доступности рибозо-5-фосфата и активности ФРДФ-синтетазы, регуляция активности которой имеет большее физиологическое значение, чем регуляция активности ФРДФ-амидотрансферазы.
Аденилосукцинатсинтетаза и IMP-дегидрогеназа являютсярегуляторными ферментами на пути превращения IMP в AMФ и ГМФ. AMФ снижает превращение IMP в аденилосукцинат, а ГМФ — превращение IMP в ксантозин –5 монофосфат, поэтому от них зависит сбалансированное содержание адениловых и гуаниловых нуклеотидов. Превращение IMP в аденилосукцинат в ходе синтеза AMФ требует ГТФ, а превращение ксантозин –5 монофосфата в ГМФ - АТФ. Перекрестная регуляция между путями использования IMP служит для того, чтобы снизить синтез одного пуринового нуклеотида при дефиците другого.
Пуриновые нуклеотиды синтезируются «запасньм путем» из азотистых оснований и нуклеозидов. Этот путьимеет вспомогательное значение, давая от 10 до 20% общего количества нуклеотидов. При этом используются азотистые основания, образующиеся процессе катаболизма нуклеиновых кислот(«путь спасения»). Два фермента катализируют эти реакции:
i)аденинфосфорибозилтрансфераза, отвечающая и образованиеAMФ из аденина иФРДФ:
Аденин +ФРДФ ® AMP +Н4Р2О7
i)гипоксантингуанинфосфорибозилтрансфераза, которая использует в качестве субстратов гипоксантин и гуанин:
Гуанин +ФРДФ ® ГМФ
Гипоксантин + ФРДФ® IMP
В результате совместного действия этих ферментов снижается выход конечного продукта обмена пуринов - мочевой кислоты.
Другой «запасной путь» включает фосфорилирование пуриновых нуклеозидов с помощью АТР. Так, аденозинкиназа катализирует фосфорилирование аденозина до AMФ или дезоксиаденозина до dAMФ:
Аденозин + АТФ ® AMФ + AДФ
Катаболизм пуриновых нуклеотидов приводит к образованиюмочевой кислоты. Мононуклеотиды теряют фосфатный остаток гидролитически под действием фосфатаз или нуклеотидаз с образованием нуклеозидов.
Дальнейшиепревращения аденозина и иозинав мочевую кислоту показаны на рисунке

Фермент аденозиндезаминазаиногда образуется в дефектной мутантной форме, что приводит к врожденному иммунодефициту, так как нуклеотиды являются регуляторами функций лейкоцитов.
Последним ферментом, участвующим в превращении азотистых оснований в мочевую кислоту, являетсяксантиноксидаза. Она окисляет гипоксантин в ксантин и ксантин в мочевую кислоту с участием молекулярного кислорода. У человека мочевая кислота является конечным продуктом обмена и выводится из организма с мочой.
Мочевая кислота является одним из нормальных компонентов мочи. За сутки в организме образуется около 1 грамма мочевой кислоты. Мочевая кислота выводится из организма с мочой - это обычный ее компонент, но в почках организма человека происходит ее интенсивная реабсорбция. Концентрация мочевой кислоты в крови поддерживается на постоянном уровне 0.12-0.30 ммоль/л.
Функции мочевой кислоты: 1. Является мощным стимулятором центральной нервной системы, ингибируя фосфодиэстеразу, которая служит посредником действия гормонов адреналина и норадреналина. Мочевая кислота пролонгирует (продлевает) действие этих гормонов на ЦНС; 2. Обладает антиоксидантными свойствами - способна взаимодействовать со свободными радикалами.
Уровень мочевой кислоты в организме контролируется на генетическом уровне. Для людей с высоким уровнем мочевой кислоты характерен повышенный жизненный тонус
Полиморфизм ферментов, участвующих в синтезе пуриновых нуклеотидов de novo, сопровождается образованием белков с низкой ферментативной активностью или нечувствительных к действию аллостерических эффекторов. При этом нарушается регуляция синтеза пуриновых нуклеотидов по механизму отрицательной обратной связи. Избыточно синтезирующиеся нуклеотиды подвергаются катаболизму, и образование мочевой кислоты повышается. Тот же результат получается при снижении активности «путей спасения» пуринов. Аденин, гуанин и гипоксантин не используются повторно, превращаются в мочевую кислоту, и возникаетгипер-урикемия. Гиперурикемия – это состояние организма, при котором содержание мочевой кислоты в сыворотке крови превышает уровень растворимости.
Сама мочевая кислота и, особенно, ее соли ураты (натриевые соли мочевой кислоты) плохо растворимы в воде. Даже при незначительном повышении концентрации они начинают начинают выпадать в осадок и кристаллизоваться, образуя камни. Кристаллы воспринимаются организмом как чужеродный объект. В суставах они фагоцитируются макрофагами, сами клетки при этом разрушаются, из них освобождаются гидролитические ферменты. Это приводит к воспалительной реакции, сопровождающейся сильнейшими болями в суставах. Такое заболевание называется подагра. Другое заболевание, при котором кристаллы уратов откладываются в почечной лоханке или в мочевом пузыре, известно как мочекаменная болезнь.
Наследственное нарушение пути реутилизации пуринов вызывает развитие у мальчиковсиндрома Леша—Нихена, при котором отмечается резкое снижение активности гипоксантингуанинфосфорибо-зилтрансферазы или она полностью отсутствует. Эта патология сопровождается 3—6-кратной гиперпродукцией уратов, образованием камней в почках, умственной отсталостью, агрессивным поведением и нанесением себе увечий.
Синтез и катаболизм пиримидиновых оснований (рис.9)
Синтез УМФ de novo, включающий 6 последовательных стадий, протекает главным образом в цитозоле клеток при участии 3 ферментов, 2 из которых полифункциональны:
- первый полифункциональный фермент содержит домены, имеющие активностькарбамоилфосфат-синтетазы II (КФС II),аспартаттранскарбамоилазы (АТК) идигидрооротазы и катализирующие 3 первые реакции этого метаболического пути;

Рисунок 9
- митохондриальнаяNAD-зависимая дегидрооротатдегидрогеназа окисляет дигидрооротат в оротат;
- превращение азотистого основания оротата в нуклеотид и его последующее декарбоксилирование до УМФ катализирует второй полифункциональный фермент, обнаруживающийоротатфосфорибозилтрансферазную и оротатдекарбоксилазную активность.
НМФ- иНДФ-киназы катализируют превращение НМФ в полифосфорные производные за счет переноса фосфата АТФ на НМФ с образованием НДФ и НТФ.
СТР-синтаза (ЦТФ- синтаза) превращает УТФ в ЦТФ, используя амидную группу глутамина и энергию АТР для аминирования пиримидинового кольца.
Аллостерически регулируется активность 2 ферментов первого полифункционального комплекса:
КФС II ингибируется УТФ и пуриннуклеотидами, но активируется ФРДФ, а АТК ингибируется ЦТФ, но активируется АТФ. Количество обоих полифункциональных ферментов регулируется также на генетическом уровне путем репрессии и дерепрессии.
Подобно пуринам, пиримидиновые азотистые основания и нуклеозиды могут превращаться в нук-леотиды«запасными путями» в реакциях, катализируемыхпиримидинфосфорибозилтрансферазой и ури-динцитидинкиназой.
Синтез пуриновых и пиримидиновых нуклеотидов строго координируется: ФРДФ активирует оба синтеза, а накопление пурин - и пиримидиннуклеотидов ингибирует образование ФРДФ по механизму отрицательной обратной связи.
Описано редкое наследственное заболевание — оротацидурия, при котором в результате мутации в гене второго полифункционального фермента нарушается превращение оротата в УМФ. С мочой выделяется до 1,5 г оротата (в 1000 раз больше, чем в норме), и развивается недостаточность пиримидиновых нуклеотидов.

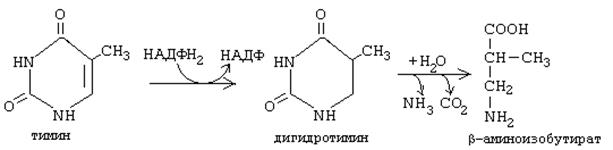
Ферментные системы организма способны разрушать пиримидиновые основания с образованием продуктов. Цитозин распадается с образованием CO2, NH3 и b-аланина.
Тимин распадается подобно урацилу, но сохраняется CH3-группа, и вместо b-аланина образуется b-аминоизобутират, а также CO2 и NH3.
Поскольку тимин встречается только в ДНК, то по уровню b-аминоизобутирата в моче судят об интенсивности распада ДНК.
b-аланин хорошо растворим в воде, имеет физиологическое значение, содержится в плазме в свободном виде или включается в мышечные белки - карнозин и ансерин.
NH3 используется для синтеза мочевины.
|
из
5.00
|
Обсуждение в статье: Синтез и катаболизм пуриновых оснований |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы