 |
Главная |


СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА
|
из
5.00
|
Половыми клетками являются мужская половая клетка сперматозоиди женская половая клетка — яйцеклетка. Половые клетки гаплоидны.
СПЕРМАТОЗОИД.Состоит из головки и хвостика и имеет длину 60—70 мкм. В свою очередь, в хвостике различают связующий, промежуточный, главныйи дистальныйотделы.
На обычных гистологических препаратах ядра сперматозоидов обладают резко выраженной базофилией и имеют вытянутую грушевидную форму (рис. 5.1). Ядро характеризуется плотным расположением хроматина, в котором наряду с гистоновыми находятся также простые негистоновые белки, обладающие положительным зарядом. ДНК в хроматине упакована параллельно, поэтому ядро имеет кристаллоподобпую структуру, его объем сведен до минимума. Ядро сперматозода гаплоидно, содержит 22 аутосомы и 1 половую хромосому, которая может быть либо Х-, либо Y-хромосомой. Количество сперматозоидов с Х- или Y-хромосомой примерно одинаковое. Ядерная оболочка сперматозоидов полностью лишена ядерных пор.
В передней части ядра под цитолеммой сперматозоида находится ак-росома— производное комплекса Гольджи и аналог лизосомы. Ее мембрана спереди прилежит к цитолемме, а сзади — к ядерной мембране. В акро-соме содержатся ферменты (10—12 различных ферментов), расщепляющие компоненты прозрачной оболочки яйцеклетки: гиалуронидаза, протеазы, гликозидаза, липаза, нейраминидаза, фосфатазы и др.
Цитоплазма сперматозоида редуцирована до минимума и очень тонким слоем покрывает ядро.
Связующий отдел хвостика содержит проксимальную центриоль. которая прилежит к ядру, располагаясь в углублении ядерной оболочки. Здесь же находится дис-тальная центриоль. От нее отходи! осевая нить — аксонема,имеющая структуру реснички и состоящая ил 9 периферических дуплетов микротрубочек и двух расположенных в центре одиночных микротрубочек Аксонема продолжается во все отделы хвостика, редуцируясь в дис-талыюм отделе. Снаружи напротив каждого дуплета дистальной цент-риоли в связующем отделе находится одна так называемая сегментированная колонна(следовательно, всего количество сегментированных колони равно 9).
 В промежуточном отделе хвостика сегментированные колонны продолжаются в 9 плотных волокон.В этом же отделе вокруг аксонемы и плотных волокон в виде спирали располагаются митохондрии. В главной части хвостика два из девяти плотных волокон, расположенные напротив друг друга, резко утолщаются и превращаются в продольные столбы,которые соединяются между собой боковыми ребрами.Формируется внешняя волокнистая оболочка,придающая жесткость и упругость хвостику. Таким образом, сегментированные колонны, плотные волокна, продольные столбы и соединяющие их ребра, а также внешняя волокнистая оболочка образуют каркас хвостика. В дистальном отделе хвостика количество микротрубочек сильно редуцируется. Снаружи хвостик покрыт цитолеммой. Благодаря движению хвостика сперматозоиды могут двигаться со скоростью 1—5 мм в минуту. Электронномикроскопическое строение сперматозоида отражено на рис. 5.2.
В промежуточном отделе хвостика сегментированные колонны продолжаются в 9 плотных волокон.В этом же отделе вокруг аксонемы и плотных волокон в виде спирали располагаются митохондрии. В главной части хвостика два из девяти плотных волокон, расположенные напротив друг друга, резко утолщаются и превращаются в продольные столбы,которые соединяются между собой боковыми ребрами.Формируется внешняя волокнистая оболочка,придающая жесткость и упругость хвостику. Таким образом, сегментированные колонны, плотные волокна, продольные столбы и соединяющие их ребра, а также внешняя волокнистая оболочка образуют каркас хвостика. В дистальном отделе хвостика количество микротрубочек сильно редуцируется. Снаружи хвостик покрыт цитолеммой. Благодаря движению хвостика сперматозоиды могут двигаться со скоростью 1—5 мм в минуту. Электронномикроскопическое строение сперматозоида отражено на рис. 5.2.

ЯЙЦЕКЛЕТКА.Это женская половая клетка с гаплоидным набором хромосом. В процессе овуляции из яичника выходит овоцит второго порядка с незавершенным мейозом, заблокированным на метафазе 2 мейоти-ческого деления. Этот блок хромосомного аппарата овулировавших овоцитов 2 порядка достаточно стабильный, так что клетки могут длительно сохраняться в метафазе II мейоза. Блокада мейоза снимается лишь при оплодотворении, и завершение мейоза с образованием зрелой яйцеклетки происходит сразу после него.
Яйцеклетка (оотида)человека имеет округлую форму (рис. 5.3, 5.4). Ее диаметр равен около 130 мкм. Содержит 23 хромосомы, одна из кото-рых половая Х-хромосома. При электронномикроскопическом исследовании в цитоплазме яп цеклетки выделяются митохонд рии, комплекс Гольджи, хорошо развитые гранулярная и аг-ранулярная эндоплазматическая сеть, а также включения: трофические (гранулы вителлина, липидные) и пигментные. Снаружи яйцеклетка окружена плазматической мембраной. Под ней располагается толстый слой цитоплазмы толщиной 2— 3 мкм. Вместе с цитолеммой его называют кортикальным слоем.В кортикальном слое находятся кортикальные гранулы,содержащие различные ферменты, в том числе и овопероксидазу,действие которой на блестящую оболочку после оплодотворения резко изменяет свойства последней. Кортикальный слой играет важную роль в организации яйцеклетки, а также в оплодотворении.
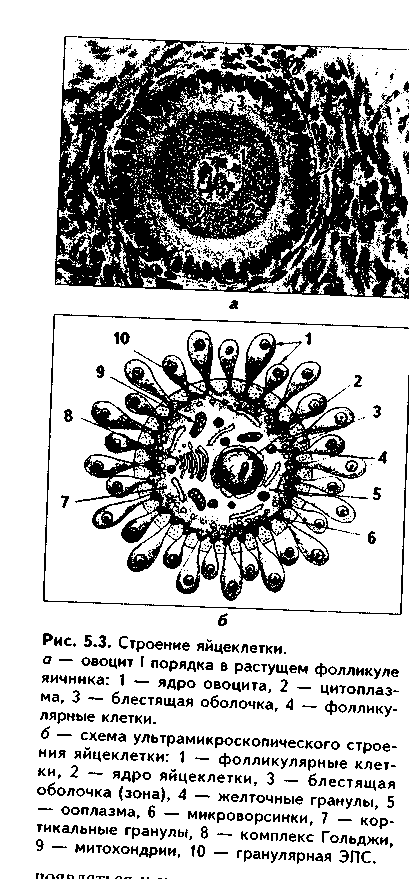
|
Яйцеклетки имеют хорошо развитый и своеобразно организованный цитоскелет. Его компоненты связаны с цитолеммой и вызывают постоянную модификацию поверхности клетки, в которой могут
появляться и исчезать микроворсинки, меняться локализация рецепторов. Яйцеклетки окружены блестящей оболочкой и слоем фолликулярных клеток. Между блестящей зоной и цитолеммой овоцита имеется небольшое перивителлиновоепространство, которое существенно увеличивается после оплодотворения, поскольку в него выделяются содержимое кортикальных гранул. Фолликулярные клетки через отверстия в блестящей оболочке посылают к цитолемме овоцита свои отростки. В ряду позвоночных в зависимости от наличия желтка, его количества и распределения по ооплазме различают алицетальные(без желтка) и леци-тальные(с желтком) яйцеклетки.

Лецитальные клетки делятся на олиголецитальные(маложелтковые) и полилецитальные(многожелтковые).
В зависимости от распределения желтка по ооплазме яйцеклетки делятся на изолецитальные, умеренно телолецитальные и резко телолецитальные. В изолецитальных яйцеклетках желток распределен равномерно. В уме ренно телолецитальных яйцеклетках желток находится на одном полюсе, который называется вегетативным. На другом полюсе — анимальном — лс жат органеллы и ядро. В резко телолецитальных клетках вегетативный но люс выражен особенно сильно и занимает подавляющую часть клетки.
Яйцеклетки млекопитающих, в том числе и человека, являются олиго-изолецитальными: содержат очень мало желтка, который равномерно рас пределен по ооплазме.
РОЛЬ ЯДРА И ЦИТОПЛАЗМЫ В ПЕРЕДАЧЕ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ
Наследственная информация хранится в ядре половых клеток, в хромосомах. Она закодирована в триплетах ДНК — генах. Ядро половых клеток выполняет три основные функции, присущие ему и в других клетках:
1. Хранение и передача наследственной информации.
2. Реализация наследственной информации.
3. Контроль за синтетическими процессами, осуществляемыми в цитоплазме.
Первая функция обеспечивается путем репликации молекул ДНК. Это позволяет при митозе двум дочерним клеткам получать одинаковые объемы генетической информации. Для сохранения генетической информации в неизмененном виде в ядре имеются ферменты, ликвидирующие повреждения молекул ДНК. Генотип половых клеток содержит только половину программы развития, строения и функционирования индивидуума, полная программа создается и реализуется только после оплодотворения.
Вторая и третья функции обеспечиваются процессами транскрипции на молекулах ДНК различных информационных, транспортных и рибосо-мальных РНК. В ядре происходит также образование субъединиц рибосом путем соединения синтезированных в ядрышке рибосомных РНК с рибо-сомными белками. Эти белки синтезируются в цитоплазме и переносятся в ядро. Субъединицы рибосом выходят в цитоплазму и соединяются с информационной РНК. После этого происходит синтез белков, необходимых для дифференцировки и специализации, а также выполнения клетками функций. Кроме ядра, некоторое количество ДНК содержится в цитоплазме в митохондриях, которые могут самостоятельно синтезировать определенные белки для собственных нужд. В цитоплазме вырабатываются вещества (триггерные белки), способные проникать в ядро и регулировать активность генома клеток. Таким образом, цитоплазма и ядро тесно взаимосвязаны is выполнении функций по хранению и передаче наследственной информации. Итак, программа эмбрионального развития индивидуума заключена в ДНКзиготы и реализуется на основе экспрессии и репрессии генов, дифференцировки клеток под влиянием зпигеномных стимулов и стимулов микроокружения.
ПРОГЕНЕЗ
Прогенезомназывается процесс образования половых клеток, или га-метогенез.В свою очередь, гаметогенез делится на сперматогенез(образование сперматозоидов) и овогенез(образование яйцеклеток). Развитие половых клеток в эмбриогенезе человека начинается довольно рано. Они возникают во внезародышевой желточной энтодерме в конце 3-й недели эмбриогенеза. Позднее эти клетки (они называются гонобластами) мигрируют в закладку половых желез на медиальной поверхности первичной почки и принимают участие в образовании половых желез — гонад.
СПЕРМАТОГЕНЕЗ.Процесс развития мужских половых клеток — сперматозоидов — называется сперматогенезом. В сперматогенезе различают 4 фазы: размножения, роста, созреванияи формирования(рис 5.5).
На протяжении периода размножения мужские половые клетки представлены сперматогониями.Это мелкие округлые клетки, делящиеся митозом. Они подразделяются на темныеи светлыесперматогонии. Темные сперматогонии являются истинными стволовыми клетками, устойчивы к действию вредных факторов и способны совершать редкие митотические деления. Светлые сперматогонии подразделяются на А и В-сперматогонии. А-снерматогонии являются полустволовыми клетками, способными к частым митотическим делениям. При делении каждой такой сперматогонии могут возникать либо две А-сиерматогонии, либо одна А- и одна В-спер-матогопия. В-снерматогонии также способны митотически делиться, но при этом не происходит цитотомии, и клетки оказываются связанными между собой цитоилазматическими мостиками. При этом возникают клоны (ассоциации)клеток.
После некоторой паузы В-сперматогонии вступают в период роста, в течение которого превращаются в сперматоциты первого порядка.Для периода роста характерен значительный рост ядра и цитоплазмы развивающихся клеток, их размеры увеличиваются в четыре и более раз.
Сперматоциты I порядка митотически не делятся, но вступают в период созревания, который состоит из двух последовательных делений мейоза (мейоз I и мейоз II).

|
МейозI называется редукционнымделением, так как в ходе его происходит уменьшение в два раза количества хромосом с формированием гаплоидного генома. Мейоз I имеет сложную профазу, состоящую из 5 стадий: лептотены, зиготены, пахитены,диплотены и диакинеза
В стадию лептотены хромосомы спирализуются и становятся видны в виде длинных тонких нитей. В стадию зиготеныгомологичные хромосомы тесно сливаются друг с другом. В это время в них происходит кросинговер — обмен генами. В стадию пахитеныхромосомы сильно спирализуются и укорачиваются. Диплотенахарактеризуется расщеплением хромосом на хроматиды и образованием тетрад. В диакинезехромосомы еще более утолщаются и несколько отходят друг от друга. Таким образом, в профазу мейоза 1 происходит подготовка к редукции
числа хромосом.
В метафазу гомологичные нары хромосом располагаются в экваторе клетки. Это ключевой момент подготовки к редукции хромосом.
В анафазу к полюсам отходят целые гомологичные хромосомы, и этот момент является началом редукции генома.
В телофазу происходит цитотомия, в результате чего образуются два сперматоцита II порядкас гаплоидным набором хромосом. Их хромосомы состоят из двух хроматид.
Второе деление мейоза (мейоз II) называется эквационным,или уравнительным.Оно начинается сразу после мейоза I и протекает по типу обычного митоза. В анафазу мейоза II к полюсам отходят хроматиды, а в результате телофазы образуются сперматиды,содержащие вместо хромосом хроматиды. Сперматиды, как и сперматоциты второго порядка, содержат гаплоидный набор хромосом, каждая из которых представлена одной
хроматидой.
Все образующиеся в процессе сперматогенеза клетки (В-сперматого-нии, сперматоциты I и II порядка, а также сперматиды) остаются связанными между собой цитоплазматическими мостиками в клеточные ассоциации,или клоны.Окончательное разделение клеток происходит в фазу формирования. Сохранение цитоплазматических мостиков между клетками имеет очень большой биологический смысл. Оказывается, что для полноценной дифференцировки сперматозоидов необходим весь диплоидный геном и продукты его деятельности. Во-первых, потому, что в исходном диплоидном геноме могут содержаться дефектные, летальные аллели генов, и клетка, получившая их, погибнет, если не будет обеспечена продуктами нормального аллеля, находящегося в ядрах клеток, его получивших. Во-вторых, как известно, одни мужские половые клетки получают Х-, другие — Y- половую хромосому. Каждая из них содержит много важных генов, необходимых для развития сперматозоидов. Поэтому благодаря цитоплаз-матическим мостикам развивающиеся мужские половые клетки получают продукты деятельности диплоидного генома.
Фаза формирования является самой продолжительной фазой сперматогенеза. В процессе ее из сперматид образуются сперматозоиды.Часто эту фазу называют спермиогенезом.Она длится дольше всех остальных фаз (около 50 суток). Процесс спермиогенеза начинается с образованием из комплекса Гольджи вначале акробласта, а затем акросомы,которая содержит ферменты для разрушения яйцевых оболочек. Центросома, состоящая из двух центриолей, перемещается в противоположный полюс. Проксимальная центриоль прилежит к ядру, а дистальная делится на две части. Из одной части образуется жгутик, который превращается в осевую нить хвостика. Вторая часть играет роль базального тельца. Образуются эле-менты нитоскелета: сегментир<| ванные колонны, плотные волок на, продольные столбы с ребрами. Цитоплазма сперматозоид.! сильно редуцируется, а ядро ста новится вытянутым, компактным и гипербазофильным. На заключительных этапах формирования сперматозоиды отделяются от соединяющей их друг с другом общей цитоплазмы и становятся свободными. Оставшийся после отделения объем цитоплазмы (остаточные тельца)подвергается фагоцитозу.

ОВОГЕНЕЗ(рис. 5.6) Принципиально протекает сходно со сперматогенезом, но имеет ряд отличий. Исходными клетками в овогенезе являются первичные половые клетки (гоноблас-ты),развивающиеся в раннем эмбриональном периоде в женской половой железе — яичнике. Эти клетки входят в состав эпителия индифферентной половой железы.В дальнейшем данный эпителий вырастает в виде тяжей в мезенхиму первичной почки (мезонефроса),а затем распадается на отдельные островки (шары Пфлюгера).В составе этих островков находятся половые клетки и окружающие их эпителиоциты (в дальнейшем - фолликулярные клетки).Гонобласты превращаются в овогонии.Эти мелкие клетки вступают в фазу размножения и интенсивно делятся митозом. В результате к концу эмбрионального развития их число достигает 7 млн. К моменту рождения период размножения заканчивается. Начиная с конца 3-го месяца эмбриогенеза и до рождения девочки одни овогонии превращаются в овоцит I порядка,другие же продолжают делиться. После рождения размножение овогонии прекращается, все они превращаются в овоциты I порядка, которые блокируются на стадии диплотены первого мейотического деления.
Далее овоциты I порядка вступают в длительный период роста. Период роста делится на две части: период малого,или медленного,роста (длится от рождения до полового созревания), и период большого,или быстрого,роста (происходит циклически на протяжении каждого менструального цикла). В период быстрого роста идет подготовка к мейозу. Таким образом, период роста может составлять 12—50 лет. Третья фаза овогенеза — созревание — начинается перед овуляцией. Происходит первое мейотическое деление, образуется овоцит II порядка, который вступает во второе мейотическое деление, но блокируется в метафазе, а также редукционное тельце.Завершение созревания инициируется оплодотворением. В отличие от сперматогенеза возникающие в результате двух делений мейоза клетки не равны по размерам. Из овоцита I порядка образуется крупный овоцит II порядка и очень мелкое редукционное тельце,которое может делиться на два редукционных тельца. Из овоцита II порядка образуется оотида и третье редукционное тельце. Следовательно, в результате двух делений образуется одна яйцеклетка и три редукционных тельца, которые вскоре погибают и фагоцитируются другими клетками. Яйцеклетка теряет центриоли.

ОПЛОДОТВОРЕНИЕ. Оплодотворение — это процесс слияния мужской и женской половых клеток, который приводит к образованию одноклеточного зародыша — зиготы.Происходит в ампулярной части яйцевода. Перед оплодотворением сперматозоиды активируются под влиянием слизистого секрета яйцевода. Это явление называется капацита-цией.Во время капацитации происходят существенные изменения белковых компонентов цитолеммы спермиев: некоторые вещества удаляются, други белки существенно модифицируются. Эти изменения играют важную ро.ь для последующей акросомной реакции.Одновременно сперматозоиды ги перактивируются, их двигательная способность резко возрастает, что способ ствует их проникновению через лучистый венец и блестящую оболочку.
Для оплодотворения необходимо около 200 миллионов сперматозоидов. В эякуляте человека содержится около 300 млн сперматозоидов, которые сохраняют способность к оплодотворению в течение 2 суток. Из них только около 200 достигают воронковой части яйцеводов, где происходи! оплодотворение. При низком содержании сперматозоидов в эякуляте (так называемая олигозооспермия)из-за недостаточной литической активности оплодотворения не просходит.
Очевидно, определенную роль в сближении сперматозоидов и яйцеклетки имеет хемотаксис. По предложению М. Гартмана (1940), вещества, продуцируемые яйцеклеткой и сперматозоидом или содержащиеся в поверхностных слоях цитоплазмы и влияющие на подвижность сперматозоидов, названы гамонами(гормонами гамет). При этом сперматозоиды вырабатывают хемотаксические вещества адрогамоны,а яйцеклетка — гинога-моны.Яйцеклетка содержит гиногамоны 1 и 2. Гиногамон 1 активирует движение спермиев, а гиногамон 2, или фертилизин,агглютинирует их. Андрогамон 1 спермиев блокирует их движение, а андрогамон 2 растворяет оболочку яйца. В последнее время, однако, путем наблюдения за половыми клетками при экстракорпоральном оплодотворении установлено, что никакого хемотаксиса при оплодотворении не существует. Встреча сперматозоида и яйцеклетки происходит случайно, но возникшее кратковременное взаимодействие между ними приводит к очень прочным межклеточным контактам.
Сперматозоиды достигают яйцеклетки через 2 часа после осеменения и окружают ее. За счет синхронного движения жгутиков сперматозоидов яйцеклетка начинает совершать вращательные движения. После вступления в контакт с фолликулярными клетками лучистого венца наступает акросомная реакция— выделение из акросом сперматозоидов ферментов. Морфологическим проявлением акросомной реакции является слияние акросомной мембраны с цитолеммой спермия в передней части головки. Для акросомной реакции большую роль играет быстрое поступление внутрь головки спермия ионов кальция, который запускает синтез циклических нуклеотидов и повышает активность АТФазы. Это приводит к увеличению внутриклеточного рН и включению акросомной реакции.
Из ферментов акросомы наибольшее значение имеют гиалуропидаза и трипсиноподобный фермент акрозин.Они воздействуют на лучистый венец и разрыхляют его: расщепляют связи между клетками, в результате чего последние диссоциируют, создавая возможность проникновения спер-миев к блестящей зоне. Важную |«>.н. играет также денудация яйцеклетки и яйцеводах — частичное или даже полное освобождение яйцеклетки от кле-iок лучистого венца. При полной денудации епермии сразу взаимодействуют с блестящей зоной.
Блестящая зона является более существенным барьером на пути сперматозоидов. Вначале спермин связываются со специфическими рецепторами на блестящей зоне. Наиболее известным рецепторным белком для сперматозоидов является гликопротеид ZP3(аббревиатура от zona pellucida — блестящая зона, оболочка). Дополнительнм рецептором является ZP2.Прикрепление сперматозоидов к блестящей оболочке является видоспецифическим. После прикрепления к блестящей оболочке спермия ферменты, связанные с внутренней акросомной мембраной, растворяют тот небольшой участок зоны, к которому прикрепился спермий. Активные движения хвостика позволяют сперматозоиду мигрировать через блестящую оболочку за 5-10 мин. Касание одного из сперматозоидов оолеммы яйцеклетки приводит к образованию на поверхности воспринимающего бугорка.Оолемма содержит систему рецепторов для взаимодействия с рецепторами спермия. После взаимодействия и слияния плазматических мембран спермия и яйцеклетки мембраны спермия включаются в состав ооплазматической мембраны, а головка спермия внедряется в ооплазму.
МЕХАНИЗМЫ БЛОКАДЫ ПОЛИСПЕРМИИ.Несмотря на прикрепление к яйцеклетке одновременно большого числа спермиев, только один из них вносит в нее свой геном. В случае проникновения ядер нескольких сперматозоидов (полиспермия)формировались бы дополнительные веретена деления, что привело бы к аномальному расхождению хромосом при дроблении, формированию недиплоидных зигот и прекращению дальнейшего развития зародыша. Для предотвращения полиспермии существует несколько механизмов.
1. Одновременно с началом взаимодействия двух гамет в яйцеклетке происходит кортикальная реакция, запускаемая быстрым повышением в яйцеклетке концентрации ионов кальция. При этом кортикальные гранулы быстро перемещаются под цитолемму и их содержимое выделяется в перивителлиновое пространствопод блестящую оболочку. В результате из блестящей оболочки формируется твердая оболочка оплодотворения,непреодолимая для спермиев.
2. Кортикальные гранулы содержат ферменты, в том числе различные гидролазы. Эти ферменты расщепляют рецепторы ZP2 и модифицируют ZP3 блестящей оболочки, которая при этом теряет способность связывать другие спермин. Это препятствует развитию полиспермии. Одновременно содержимое кортикальных гранул блокирует акросомную реакцию в других спермиях. Все эти опосредованные через блестящую оболочку изменения обеспечивают позднюю блокадуполиспермии. 3. Кроме вышеизложенного, кортикальные гранулы изменяют молеку лярную организацию оолеммы, которая приобретает новые свойства, в том числе и отрицательный заряд, отталкивающий отрицательно заряженныг спермин (ранний блок полиспермии).
СИНКАРИОН. Ядра сперматозоида и яйцеклетки (с этого момент;! они называются мужским и женским пронуклеусами) набухают, в них об наруживаются ядрышки (рис. 5.7). Набухание мужского пронуклеуса про
исходит вследствие замены и хроматине протаминов на гис-тоны. Пронуклеусы приближа ются друг к другу, терякл ядерные оболочки и сливаются. Процесс слияния пронукле-усов называется синкарионом. При этом их геномы перемешиваются, и восстанавливается диплоидный набор хромосом. В результате образуется новый организм — зигота.
ИЗМЕНЕНИЯ, ПРОИСХОДЯЩИЕ В ЯЙЦЕКЛЕТКЕ ПОСЛЕ ОПЛОДОТВОРЕНИЯ. 1. В первые 10 мин. после оплодотворения в зиготе усиливается углеводный обмен, активируется распад гликогена, что свидетельствует о потреблении энергии. 2. В большинстве случаев резко повышается потребление кислорода. 3. В первые минуты увеличивается содержание нуклеиновых кислот, что является признаком усиления диссимиляционных процессов. 4. Резко (в 100 и более раз) возрастает обмен фосфатов, в 10 и более раз — калиевый и кальциевый обмен. 5. Резко возрастает проницаемость мембраны для фосфатов, изменяются ее электрические свой-ства. 6. Повышается активность протеолитических ферментов. 7. Запускается синтез ДНК, и-РНК и белка.
После короткого периода покоя зигота вступает в новый период эмбриогенеза — дробление.

|
Искусственная инсеминация. Достижения современной эмбриологии позволяют решать целый ряд практических вопросов, связанных с женским и мужским бесплодием, исправления генетических дефектов. Искусственная инсеминация — это введение в половые пути женщины ранее полученной во время эякуляции семенной жидкости мужчины. Для этого эякулят замораживается в жидком азоте при температуре -196°С, где может сохраняться в течение длительного времени. В настоящее время искусственная инсеминация широко используется при мужском бесплодии (олиго- или азооспермия, т.е. существенное снижение количества полноценных активных сперматозоидов или их полное отсутствие в эякуляте). В таких случаях женщинам, желающим иметь детей, но из-за бесплодия мужа не имеющим таких возможностей, по понятным морально-этическим соображениям искусственным путем вводят в половые пути сперматозоиды мужчин-доноров. В некоторых странах благодаря технике криоконсервации создаются банки спермы. Экстракорпоральное оплодотворение, или оплодлотворение in vitro, применяют: 1. При женском бесплодии, не связанном с нарушением образования ■[ женских гамет (нарушении проходимости маточных труб). 2. После менопаузы. Применяется тогда, когда в яичнике женщины еще имеются примордиаль-ные фолликулы, но развитие их до уровня зрелых яйцеклеток и оплодотворение последних в естественных условиях невозможно. 3. Может быть использована имплантация зародыша, полученного из родительских половых клеток, в матку приемной матери (так называемое "суррогат-материнство", которое необходимо тогда, когда у "генетической" матери отсутствует или недоразвита собственная матка при полноценной функции яичников).

|
Экстракорпоральное оплодотворение включает следующие этапы: 1. Гормональная стимуляция фолликулогенеза. Применяют препараты, представляющие смесь фоллитропина и лютропина. Их введение позволяет получить в яичнике большое число синхронно развивающихся зрелых фолликулов. 2. Извлечение из яичника (под контролем ультразвукового исследования) яйцеклеток путем пункции фолликулов. 3. Оплодотворение яйцеклеток специально подготовленными сперматозоидами (размороженными или свежими). Для этого сперматозоиды отделяют от семенной жидкости путем центрифугирования, отмывания, а затем вызывают капацита-цию инкубацией в атмосфере углекислого газа. 4. Имплантация зародыша в матку женщины. Для этого зародыш вначале выращивают на питательных средах до стадии 4—8 бластомеров. Для повышения эффективности метода имплантируют не один, а несколько зародышей. При этом эндометрий матки должен быть подготовлен к имплантации. Метод экстракорпорального оплодотворения, очевидно, позволит в даль нейшсм исправлять генные аномалии: в настоящее время на эмбрионах жи вотных разработан метод микроинъекции генов в пронуклеус. После детальной диагностики генных нарушений подбираются аналогичные здоровьк гены, определяющие желательный признак. Так, на мышах проведены опы ты по инъекции в оплодотворенную яйцеклетку гена белка мышц миозина Это проявилось в сильном развитии мышц у потомства.
ДРОБЛЕНИЕ. ЗНАЧЕНИЕ И МЕХАНИЗМЫ. СТРОЕНИЕ МОРУЛЫ И БЛАСТОЦИСТЫ. ИМПЛАНТАЦИЯ
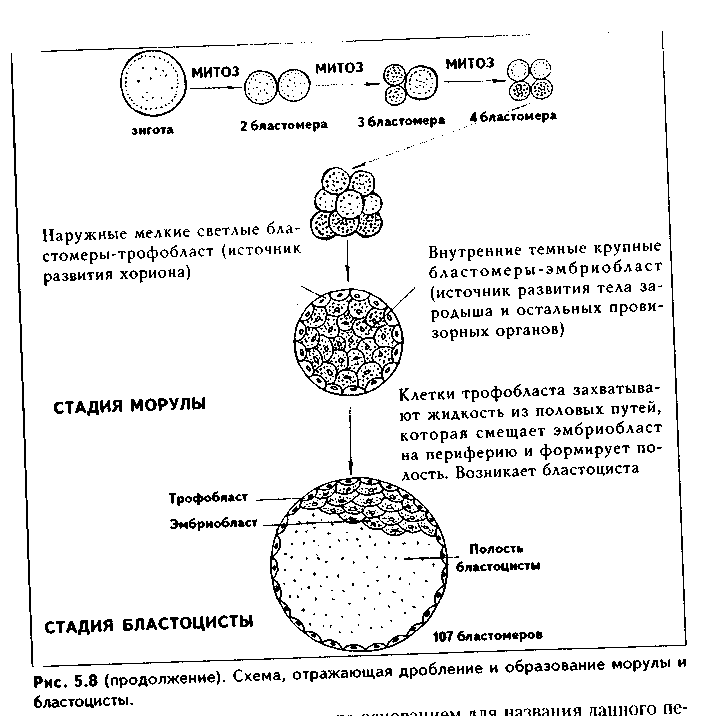
Дробление характеризуется превращением одноклеточной зиготы в многоклеточный зародыш.Это происходит в результате последовательных ми-тотических делений. При- этом из-за отсутствия С,-периода клетки не успевают увеличиваться в размерах. Поэтому с каждым делением размеры клеток уменьшаются, что послужило основанием для названия данного периода эмбриогенеза дроблением.Кроме того, общий объем зародыша в ходе дробления не только не увеличивается, а, напротив, уменьшается на 20-40%. Это свидетельствует о том, что в ходе дробления теряются какие-то вещества, и эти потери не компенсируются синтезом новых белков. Образующиеся в результате дробления клетки называются бластомерами.
Дробление зависит от количества желтка и может быть полным (го-лобластическое) и неполным (меробластичеекомомерным и неравно-
мерным, синхроннымили асинхронным.У млекч.кпающих, в том числе и у человека, дробление полное(делится весь материал зиготы), неравномерное(образуются бластомеры разной величины), асинхронное(бластомеры делятся неодновременно: за стадией двух бластомеров наступает стадия трех бластомеров, так как один из бластомеров вступает в деление позже второго, и т.д.) (рис. 5.8). Асинхронпосп. и неравномерность дробления проявляется не сразу, а начиная со второго деления, первые два бластомеры имеют одинаковые размеры.
В результате дробления образуются бластомеры разной величины крупные темные и мелкие светлые. Светлые бластомеры дробятся быстрее и быстро окружают снаружи темные бластомеры, занимающие внутреннее положение. Светлые бластомеры называются трофобластоми в дальнем! шем явятся источником развития эпителия хориона(см. ниже). Из темных бластомеров (эмбриобласт) образуются тело и провизорные органы заро дыша, за исключением хориона.
Зародыш, состоящий из плотного скопления клеток эмбриобласта и трофобласта, у млекопитающих называется морулой.Она образуется на 3-й сутки эмбриогенеза. Морулу часто отождествляют с бластулой у других животных. Между клетками морулы устанавливаются тесные межклеточные щелевые и плотные контакты (компактизация зародыша), чему способствует адгезивный белок увоморулин,встраивающийся в цитолеммы бластомеров. При этом клетки эмбриобласта связываются друг с другом при помощи щелевидных контактов (нексусов), которые обеспечивают информационные взаимодействия, тогда как в трофобдасте обнаруживаются плотные контакты, обеспечивающие его барьерные свойства. Кроме того, вплоть до стадии бластоцисты зародыш окружен блестящей оболочкой. Ее функции до оплодотворения и после него очень важны.
Функции блестящей оболочки. 1.Входя до момента овуляции в состав гемато-овариального барьера,она и в дальнейшем вплоть до своего разрушения при образовании бластоцисты выполняет барьерные функции.
2. Участвует в оплодотворении, обеспечивает его видоспецифичность, т.к. несет рецепторы к спермиям.
3. Благодаря блестящей оболочке бластомеры дробящегося зародыша располагаются компактно в ограниченном трехмерном пространстве, что играет важную роль для установления межклеточных контактов в моруле. Если удалить в это время блестящую оболочку, то компактизация нарушается, бластомеры лежат в виде цепочки, что приводит к резкому нарушению эмбриогенеза.
4. Блестящая оболочка препятствует прилипанию зародыша, клетки которого приобретают в это время выраженную адгезивпость, к слизистой яйцевода.
5. При многоплодной беременности блестящая оболочка препятствует слипанию соседних зародышей и образованию так называемых агрегационных химер.
К 4-м суткам развития клетки трофобласта начинают секретировать жидкость, которая накапливается внутри морулы и приводит к образованию полости, а также смещает эмбриобласт на один из полюсов. Так образуется бластоциста. Она состоит из бластодермы (трофобласт), бластоцеля (полость внутри) и эмбриобласта, или внутренней клеточной массы (рис. 5.8). От бластулы других животных бластоциста отличается тем, что ее клетки не однородны, а уже дифференцированы на трофобласт и эмбриобласт. При образовании бластоцисты блестящая оболочка разрушается и сбрасывается. Для того, чтобы сформировалась бластоциста, необходимо сочетание двух процессов: выработки бластомерами жидкости и создания прочной стенки зародыша. Последнее условие обеспечивается плотными контактами между клетками трофобласта. Его клетки принимают полигональную форму, в них соответствующим образом ориентируется цитоскелет и организуются цитолеммы.

|
Дробление у человека происходит в течение первой недели эмбриогенеза. За это время зародыш попадает в полость тела матки и начинает имплантироваться.
Имплантация — процесс проникновения зародыша в слизистую оболочку стенки матки (эндометрий) и установления тесных связей с ее кровеносными сосудами. Состоит из двух фаз: адгезии, или прилипания трофобласта к слизистой оболочке матки, и инвазии. Перед имплантацией трофобласт разделяется на два слоя: клеточный трофобласт, или цитотро-фобласт (внутренний листок), и симпластотрофобласт (синонимы: плазмодиотрофобласт, синцити-отрофобласт, синтрофобласт) — наружный листок. Обычно имплантация происходит в эндометрий задней стенки матки, причем в том его участке, где залегают достаточно крупные кровеносные сосуды.
Симпластотрофобласт выделяет ферменты, которые лизируют ткани эндометрия. За счет этого происходит инвазия — внедрение зародыша в слизистую оболочку. Трофобласт разрушает кровеносные сосуды эндометрия, из которых изливается кровь, омывая зародыш. После инвазии края слизистой оболочки над зародышем срастаются (рис. 5.9). Таким образом, у человека имплантация является глубокой, интерстициальной, поскольку зародыш глубоко проникает в эндометрий, разрушая его сосуды.
Имплантация длится около 40 часов. В ходе имплантации изменяет-ся тип питания зародыша. В течение короткого времени после онлодотвор* ния зародыш использует небольшие запасы питательных веществ, содерж.i шихся в яйцеклетке (аутотрофный тип питания). После расходования m;i териала желточных включений зародыш переходит на гистотрофныйтип питания, используя секрет слизистых клеток эпителия яйцевода, матки. . также маточных желез, и продукты распада тканей в начальные фазы ими лантации. Наконец, после разрушения сосудов эндометрия, устанавливается гемотрофныйтип питания зародыша.
|
из
5.00
|
Обсуждение в статье: СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы