 |
Главная |


Нейронные механизмы рабочей (оперативной) памяти
|
из
5.00
|
Есть способы классификации памяти, основанные на ее функциональном значении. Оперативная память связана с хранением информации, необходимой для решения конкретной задачи, в течение времени, которое требуется для ее решения. Тем не менее она отличается от кратковременной памяти, поскольку к оперативной памяти относятся только те следы, которые были воспроизведены, тогда как в кратковременной памяти хранится и воспроизведенная, и невоспроизведенная информация. В оперативной памяти характерно сочетание сиюминутного сознания и мгновенного воспоминания о накопленной ранее информации (Гольдман-Ракич, 1992). Она имеет дело с фактами и цифрами и обеспечивает срочную активацию и кратковременное хранение символической информации, а также возможность
манипулирования ею.
Примером оперативной памяти может служить операция переноса при вычислении в уме, требующая временного запоминания ряда чисел и удержания результата одного сложения, пока подсчитывается вторая сумма. Другими примерами могут быть планирование шахматных ходов, построение предложения в разговорной речи. Кратковременная память имеет решающее значение в процессе продукции и понимания речи, при обучении и мышлении. Информация из оперативной может стать долговременной, если позднее будет прочно зафиксирована, или может исчезнуть, не оставив следа.
Факты, полученные при наблюдении за больными, имеющими повреждения префронталъной области коры больших полушарий мозга, свидетельствуют о ее участии в механизме формирования оперативной памяти. Больные с такими нарушениями испытывают большие трудности в применении имеющихся у них знаний в повседневной жизни, хотя накопленная ранее информация нередко сохраняется в полном объеме, и после травмы показатели в традиционных тестах оценки интеллектуальных способностей не меняются. Значимость префронтальной коры подтверждают и онтогенетические данные. Для оценки познавательных способностей детей используют тест постоянства объекта, разработанный Жаном Пиаже. Он представляет собой вариант теста с отсроченным ответом. Ребенку показывают две коробки, в одной из которых лежит игрушка. Коробки закрывают, ребенка намеренно отвлекают, а спустя короткое время просят показать ту, в которой находилась игрушка. После того как ребенок несколько раз правильно покажет коробку, у него на глазах игрушку перекладывают в другую и отмечают, какую коробку выберет ребенок в этом случае. Правильность ответов детей зависит от степени развития их префронталъной коры.
Дети, не достигшие восьмимесячного возраста, у которых нейрональная организация коры еще не имеет характеристик, свойственных взрослому человеку, плохо справляются с такими заданиями и продолжают выбирать ту коробку, на которую указывали раньше, но в которой уже нет игрушки. Так же плохо справляются с ними обезьяны, у которых хирургически удаляли пре-фронтальную кору. Оперированные обезьяны просто повторяли предыдущий ответ, не меняя его в связи с вновь поступившей информацией.
Ж.П.Буржуа и П. Гольдман-Ракич изучали скорость, с которой формируются связи в префронтальной коре детенышей обезьян. Они обнаружили, что быстрее всего образуются синапсы в префронтальной коре в возрасте двух-четырех месяцев, когда у обезьян появляется способность к выполнению тестов с отсроченным воспроизведением (Гольдман-Ракич, 1992).
Активность отдельных нейронов в префронтальной коре подробно изучена Дж. Фастером, К. Кубота и X. Ники. Обезьянам вводили в префронтальную область коры тонкие электроды, регистрирующие нейронную активность, и обучали выполнять простые тесты с отсроченным ответом. Выявилась четкая специфичность в ответах нейронов. Электрическая активность одних клеток усиливалась при предъявлении информации, других — только во время отсрочки, когда животные ее припоминали. Нейроны третьей группы реагировали тогда, когда начиналась двигательная реакция (Гольдман-Ракич, 1992).
Специфическая активность нейронов в процессе отсрочки показана в эксперименте, где регистрировали электрическую активность отдельных нейронов префронтальной коры в тесте с отсроченным ответом. Предварительно обезьяну обучали фиксировать взор на маленьком пятне в центре телевизионного экрана. Затем в одном из восьми участков экрана на короткое время появлялся раздражитель (обычно это небольшой квадрат). В конце отсрочки продолжительностью от трех до шести секунд центральное пятно, на котором фиксировался взор, выключалось, что служило животному сигналом о необходимости перевести взгляд в ту точку экрана, где перед отсрочкой появлялся раздражитель. В случае правильного ответа животное получало сок. Поскольку взор животного был фиксирован на центральном пятне, каждый из зрительных раздражителей активировал определенную группу клеток сетчатки. Эти клетки приводили в действие лишь определенные от- " делы зрительных проводящих путей (рис. 15.1) (Гольдман-Ракич, 1992).
С помощью этого опыта удалось показать, что некоторые нейроны префронтальной коры обладают своего рода полями памяти: когда из поля зрения животного исчезает цель, имеющая для него особую значимость, некоторые нейроны префронтальной коры начинают генерировать электрические сигналы вдвое чаще. Они остаются возбужденными до конца отсрочки, т. е. до того момента, когда животное переводит взгляд в нужную точку экрана. Каждый нейрон кодирует своей активностью определенное положение зрительного раздражителя.
Нейроны, способные хранить информацию о зрительном и пространственном расположении сигнала (отвечающие за пространственную кратковременную память), по-видимому, организованы в префронтальной коре в специфическую область. Если во время отсрочки их работа нарушается, возможны ошибки при воспоминании.
Поскольку префронтальная кора функционирует как посредник между памятью и действием, можно предположить, что повреждение этой области мозга не наносит ущерба знаниям животного о внешнем мире, но нарушает его способность осознавать эти знания и пользоваться ими. Действительно, обезьяны с поврежденной префронтальной корой, как и многие люди с аналогичными травмами, без труда обучаются выполнять тесты на
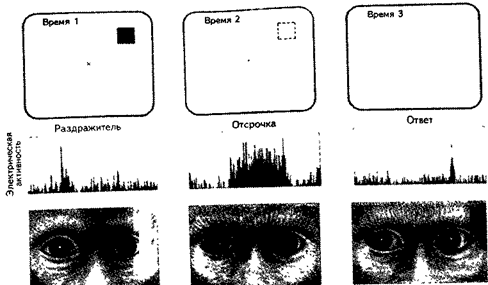
Рис. 15.1. Тест с отсроченным ответом для изучения оперативной памяти у обезьян. Когда обезьяна фиксирует взор на центральном пятне, на экране (слева) вспыхивает и затем исчезает цель. Во время длящейся несколько секунд отсрочки обезьяна хранит об этой цели “мысленную” память (в центре). Когда центральное пятно исчезает, животное переводит взгляд туда, где появлялась цель (справа). Как показывает регистрация электрической активности, некоторые нейроны префронтальной коры реагируют на появление цели, другие сохраняют о ней “мысленную” память, а третьи разряжаются, подготавливая двигательный ответ (Гольдман-Ракич, 1992).
сенсорное различение. Все формы ассоциативной, долговременной, памяти сохраняются до тех пор, пока испытуемый воспринимает знакомые внешние раздражители, ассоциированные с определенными последствиями и ожиданиями (Гольдман-Ракич, 1992).
Префронталъная кора является частью сети реципрокных связей между центральной бороздой и основными сенсорными, лимбическими и премоторны-ми зонами коры. Эта сеть отвечает за переработку пространственной информации. Нейроны в центральной борозде чувствительны также к специфическому расположению зрительных раздражителей, а значит, имеют доступ к зрительной и пространственной информации, перерабатываемой в других отделах мозга. Центральная борозда получает сигналы из задней теменной коры, где идет анализ информации, связанной с пространственным зрением.
Кратковременная память зависит от доступности информации, хранящейся в долговременной памяти. Эффективность долговременной памяти определяется сохранностью гиппокампа. Метод авторадиографии выявил, что гиппокамп и область центральной борозды часто активируются одновременно во время выполнения тестов с отсроченным ответом. П.С. Гольдман-Ракич полагает, что главная функция гиппокампа связана с консолидацией следов памяти, а префронтальной коры — с извлечением результатов такого обучения из долговременного хранилища в других отделах мозга и их непосредственным включением в оперативную деятельность.
|
Однако функционирование рабочей памяти зависит и от других структур мозга. Запись электрической активности в процессе запоминания у человека обнаружила появление медленной позитивной волны в лобно-цен-тральных областях на слова, которые впоследствии были узнаны (Friedman, 1990). Существует достаточно данных о том, что эффективность мнемичес-кой деятельности связана с развитием таламокортикальных сетей и частотой альфа-ритма (Khmesh e. a , 1990).
В префронтальной коре выявлено высокое содержание катехоламинов, прежде всего дофамина, регулирующего клеточную активность, связанную с кратковременной памятью. Нейроны некоторых слоев мозговой коры богаты определенными рецепторами дофамина — D1. Именно эти нейроны проецируются в таламус, где происходит переключение потока информации в кору.
Префронтальная кора и области, с которыми она активно связана (гиппо-камп, нижняя часть теменной коры, таламус), характеризуются высокой метаболической активностью при выполнении теста с отсроченным воспроизведением. Метаболическая активность мозга имеет разный характер — в зависимости оттого, требует ли задание от испытуемого памяти о местоположении предметов или об их признаках. При необходимости запоминать ряд слов или произносить глагол, ассоциативно связанный с высвеченным на экране существительным, отмечалось усиление работы нейронов префронтальной коры.
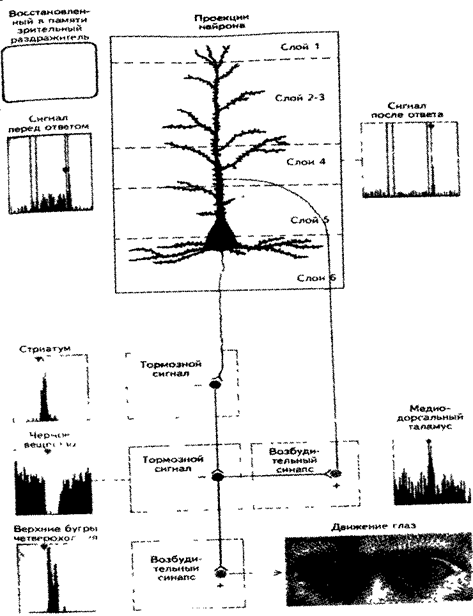
П.С. Гольдман-Ракич полагает, что префронтальная кора разделена на множество участков памяти, каждый из которых специализируется на кодировании особого рода информации, например, о местоположении, отдельных признаках объектов, таких как цвет, размеры, форма, а у людей — еще и семантических и математических знаний. Эти нейроны также выполняют функцию возбуждения или торможения других структур мозга. Кле1 -ки каждого слоя коры образуют в мозге строго специфические системы связей (рис. 15.2) Клетки одного типа, находящиеся в пятом слое коры, проецируются в хвостатое ядро, скорлупу и верхние бугры четверохолмия. Нейроны шестого слоя проецируются в таламус, через который к коре проходят сенсорные сигналы с периферии мозга
Префронтальная кора не может самостоятельно инициировать двигатель-!!
ные ответы, однако она контролирует движение, программируя, облегча^
или подавляя команды тем мозговым структурам, которые управляют сокра.д
щением или расслаблением мышц. , ^
т.
i”T
|
из
5.00
|
Обсуждение в статье: Нейронные механизмы рабочей (оперативной) памяти |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы