 |
Главная |


Тема 2. ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ ПРИЗНАКОВ
|
из
5.00
|
Цель и задачи темы. Изучить закономерности наследования родительских признаков гибриднъш потомством в первом, втором и последующих поколениях, раскрыть явление доминирования и рецессивности признаков и вероятностный характер их соотношения при расщеплении во втором и последующих гибридных поколениях моно- и полигибридных скрещиваний, показать, какие цитологические основы обусловливают расщепление признаков в гибридном потомстве, показать отличие характера числового расщепления признаков F 2 дигиб-ридного скрещивания при взаимодействии неаллельных генов, от менделевского числового соотношения 9:3:3:1 и какие закономерности наследственности вытекают из работ Менделя — дискретная природа наследственности, относительное постоянство гена и аллельное состояние гена. Проработав тему, необходимо уметь:
— пользоваться терминами: родительское поколение (Р), первое гибридное поколение ( Fi ), второе гибридное поколение (F2), доминантный, рецессивный, гомозиготный, гетерозиготный, расщепление, независимое распределение, неполное доминирование;
— дать определение генотипа и фенотипа, сравнив эти понятия и разъяснив их связь в понятиями «доминантный» и «рецессивный»;
— проиллюстрировать с помощью решетки Пеннета скрещивания по одному, двум и больше признакам и указать, какие численные соотношения генотипов и фенотипов следует ожидать в потомстве от этих скрещиваний;
— сопоставить типы наследования признаков при скрещиваниях с поведением хромосом во время мейоза и оплодотворения;
— изложить своими словами правила наследования, расщепления и независимого распределения признаков, открытых Г. Менделем-
Закономерности наследования потомками признаков организмов впервые были открыты основоположником генетики чешским ученым Грегором Менделем. Его работа, опубликованная в 1866 г. «Опыты над растительными гибридами», является классическим произведением. Она отличается необычайной силой и внутренней законченностью. Свои опыты по гибридизации гороха Мендель провел и обработал (1858—
1865 гг.) с поразительной ясностью, свойственной мышлению гения.
Следует учесть, что на протяжении столетий предшественники Менделя, изучавшие наследственность и изменчивость организмов, не сумели открыть закономерности наследования признаков, поскольку пытались суммарно по большому количеству признаков определить степень сходства и различия родителей и потомков. Суммарная оценка не могла привести к выяснению законов наследственности, так как охватить точными наблюдениями сразу большое число признаков очень трудно, к тому же разные признаки наследуются неодинаково. В отличии от своих предшественников Г. Мендель исследовал сложное явление наследственности аналитическим путем. Он разработал и применил принципиально новый метод генетического анализа наследования признаков.
Необходимо знать основные положения метода гибридологического анализа. Это и правильный выбор с полным пониманием поставленной задачи биологического объекта — гороха, растения с хорошо заметными альтернативными признаками и самоопылителя, и отбор для своих опытов только гомозиготных сортов (из 34 собранных им сортов после двухлетней проверки для опытов оставлены только 22 гомозиготных сорта), и аналитическое изучение с точным количественным индивидуальным учетом в нескольких поколениях каждого отдельного признака, а затем совместное наследование этих нескольких признаков, не принимая во внимание всех остальных признаков. Введение впервые Менделем математики и буквенной символики в биологический опыт дало ему возможность абстрагировать и обобщить конкретные результаты в виде математических формул и закономерностей.
Г. Мендель впервые доказал дискретность наследственности, заложив этим основы генетики. Важно отметить, что это открытие сделано Менделем задолго до цитологических открытий явлений митоза и мейоза и тех внутриклеточных процессов, которые при этом происходят. Мендель ввел понятие о наследственных факторах, позднее названных генами. Он показал, что наследуются не сами признаки, а наследственные факторы,' определяющие эти признаки, и что у каждого организма наследственные факторы — гены представлены парами: один аллель этой пары пришел с гаметой от отца, а второй от матери- Что половые клетки содержат от каждой аллельной пары только по одному наследственному фактору — гену.
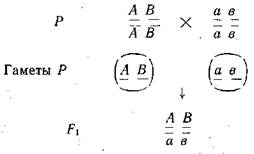
Мендель обозначил пару 'наследственных факторов парой одноименных букв. При этом наследственный фактор, определяющий доминантный признак, он обозначил заглавной буквой, а рецессивный — той же строчной буквой.
При этом следует различать такие понятия как гомозигот-ность и гетерозиготность, что особи, имеющие одинаковые фенотипы, могут иметь разные генотипы. Гомозиготными называют организмы, в соматических клетках которых одинаковые аллельные гены — АА или аа или ААВВ или ААввСС и т. п., а если е соматических клетках разные аллели генов — Аа или АаВв или АаВвСс и т. п., то их называют гетерозиготными организмами.
При изучении этой темы следует твердо запомнить установленные Г. Менделем закономерности наследования признаков — доминирование, или единообразие гибридов первого поколения, расщепление гибридов второго поколения, закон чистоты гамет, независимого комбинирования (наследования) признаков (неаллельных генов).
Следует различать типы скрещиваний: взаимные (реци-прокные), возвратные (бекроссы), анализирующие, моногиб- . ридные, полигибридные. Твердо помнить числовое соотношение генотипических и фенотипических классов во втором поколении при моногибридном, дигибридном и полигибридном скрещиваниях, а также при возвратных скрещиваниях.
Необходимо уяснить зависимость характера наследования признаков от цитологических закономерностей поведения хромосом при образовании гамет и при соединении гамет в процессе оплодотворения.
Кроме того, следует иметь в виду, что закономерности* установленные Менделем, справедливы лишь при условии, когда развитие одной пары признаков определяется парой аллельных генов и когда разные гены локализованы в разных (яегомологичных) парах хромосом и могут в результате этого свободно (независимо) комбинироваться между собой как при образовании гамет, так и при вероятном их сочетании во время оплодотворения.
Однако, как позже было установлено, многие признаки развиваются под влиянием не одной, а нескольких пар неаллельных генов, которые взаимодействуют между собой. При ! этом изменяется характер расщепления фенотипических классов и при дигибридном скрещивании не соответствует Менде-левокому соотношению 9:3:3:1.
От скрещивания двух дигетерозигот при комплементарном взаимодействии неаллельных генов может быть расщепление фенотипических классов 9:7; 9:6:1; 9:4:3, при эгоистазе— 12 : 3 : 1; 13 : 3; при полимерии — 15 : 1.
Вместе с тем, эти отличия расщепления фенотипических классов не противоречат законам наследования, установленным Г. Менделем, а наоборот, углубляют и показывают многогранность явления. При взаимодействии неаллельных генов характер передачи генов и соотношение генотйпическнх классов не изменяется-
Кроме того, следует выделить законы наследственности, вытекающие из работ Г. Менделя: закон о дискретной природе наследственности, об относительном постоянстве гена и об аллелыном состоянии его,
Для закрепления материала этой темы решите задачи 1 ... 68, приведенные в учебнике, и задачи 61 ... 83, приведенные в методических указаниях, и проанализируйте их решения.
ЛИТЕРАТУРА: 1, с. 55 ... 58; 2, с. 63 ... 81.
Вопросы для самопроверки
1. Сущность метода гибрид ологичедаого анализа, разработанного Г. Менделем.
2. Правило доминирования и единообразия гибридов первого поколения.
3. Расщепление гибридов F 2 и последующих поколений при моноги'бридном скрещивании.
4. Закон чистоты гамет.
5. Расщепление гибридов F 2 при дигибридном скрещивании-
6. Закон независимого комбинирования признаков (неаллельных генов).
7. Цитологические основы и вероятностный характер расщепления.
8. Особенности наследования признаков при взаимодействии неаллельных генов.
9. Основные законы наследственности, вытекающие из работ Г. Менделя.
Тема 3. ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
Цель и задачи темы. Показать различный характер распределения генов в половые клетки в зависимости от расло-
ложения генов в одной паре или в разных парах хромосом и характер расщепления признаков при независимом и сцепленном наследовании. Показать, что в результате кросеинговера происходит рекомбинация генов и расширяется возможность ком'бинативной изменчивости. Показать строение генетических и цитологических карт хромосом и их использование, ознакомиться с хромосомным механизмом определения пола и наследованием признаков, сцепленных с полом. Проработав тему, необходимо уметь:
— изложить своими словами суть хромосомной теории наследственности и эволюционную роль кроссинговера;
— определить группы сцепления генов;
— рассчитать процент кроссоверных особей в анализирующем скрещивании при неполном сцеплении;
— определить наследование признаков, сцепленных с по1-лом.
В первой теме было показано, что развитие экспериментальной биологии и особенно достижения цитологии во второй половине XIX и первом десятилетии нашего века создали условия для принятия и понимания открытий Г. Менделя. Открытие поведения хромосом при редукционном делении и оплодотворении показали их связи с независимым наследованием признаков, подтвердили убеждение, что самопровзводя-щиеся хромосомы являются основными носителями менделев-ских наследственных факторов — генов. Однако прямых экспериментальных данных, доказывающих локализацию генов в хромосомах, к этому времени не было.
В то же время накапливались данные, которые показывали, что некоторые признаки при размножении не менделиру-ют, а наследуются сцепленно. Явление сцепленного наследования признаков открыли в 1906 г- В. Бэтсон и Пеннет, но они не смогли раскрыть существа этого явления. Постепенно таких фактов накапливалось в опытах многих исследователей все больше и больше и до 1910 года и;х относили к исключениям из менделевского правила независимого наследования .признаков (или независимого распределения генов).
Правильное объяснение этому явлению было дано Т. Морганом и его сотрудниками в разработанной ими хромосомной теории наследственности. Они изучили в 1910—1911 гг. сцепленное наследование признаков у дрозофилы. И впервые эксперимента лыно доказали, что гены находятся в хромосомах, расположены в них линейно и образуют группы оцепления соответственно гаплоидному числу хромосом, что независимо
могут /комбинироваться и наследоваться только несцепленные гены, т. е. гены, локализованные в разных парах хромосом.
Следует отметить, что этому открытию способствовал тот редкий факт, что у самца мушки дрозофилы наблюдается-полное сцепление (полное сцепление наблюдается еще только у самок тутового шелкопряда). И в обычном дигибридном скрещивании мушки дрозофилы при скрещивании гетерозиготного по двум парам аллелыных генов самца с гомозиготной рецессивной по этим двум парам аллелей самкой в их потомстве расщепление не происходит и особей с перекомбинациеи признаков совершенно не появляется (см. в учебнике пример скрещивания особей, различающихся по окраске тела и длине крыльев).
Вместе с тем Т. Морган доказал, что сцепление очень редко бывает полным. Если в скрещиваниях дрозофилы, гетерозиготной по этим парам признаков, взять самку (у которой полного сцепления не наблюдается), а гомозиготным по рецессивным этим признакам взять самца, то в их потомстве появятся особи с четырьмя возможными комбинациями двух аллельных пар признаков (серое тело — черное тело и длинные крылья — зачаточные крылья). Однако независимого их распределения с числовым соотношением в 9:3:3: 1 не
■ происходит. А в анализирующем скрещивании особей с перекомбинированными признаками всегда меньше, чем 1/4 часть, как должно быть при независимом их распределении. Следовательно, наблюдается неполное сцепление признаков. Перекомбинация признаков происходит в результате перекреста и: обмена частями гомологичных пар хромосом при их конъюгации в профазе редукционного деления мейоза. Хромосомы, в которых произошел перекрест и обмен участками хромосом (крюосиыговер), называют кроссоверными.
Кроссинговер изменяет характер локализации генов в
..группах сцепления, происходит перекомбинация генов, расширяется возможность комбинативной изменчивости, что-очень важно для эволюции и в селекции.
Учитывая линейное расположение • генов в хромосомах,. Т. Морган объяснил частоту кроссинтовера расстоянием расположения генов в хромосоме. Чем дальше друг от друга расположены гены в хромосоме, тем чаще происходит между ними кроосинговер и тем больше образуется кроссоверных гамет (особей). За единицу измерения расстояния между изучаемыми генами принят один процент кроссинговера (морга-
нида), т. е. процент кроссоверньих особей от общего числа особей анализирующего скрещивания.
Для 'наглядности в схемах (скрещивания условные обозначения генов записывают с учетам локализации их в хромосомах. Если гены локализованы в разных хромосомах, то это показывают отдельными черточками, а если гены локализованы в одной хромосоме —■ то одной непрерывной черточкой.
Например, при независимом наследовании дигибридное скрещивание можно записать так:
Сцепленное наследование записывают следующим образом:
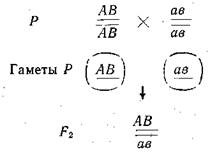
Локализация генов в одной или разных хромосомах определяет возможность образования того или иного числа типов гамет.
Так, у дигетерозиготы с локализацией генов в разных хромосомах, т. е. со свободным комбинированием, может образоваться с одинаковой частотой вероятности четыре возможных типа гамет, а при полном сцеплении — только два
родительских типа, и при неполном сцеплении — четыре возможных типа, но доля некроссоверных гамет всегда больше 50%, а доля кроссоверных — всегда меньше 50%. Например:

Изучая этот раздел, следует внимательно рассмотреть механизм определения пола и наследования признаков, сцепленных с полом, поскольку это были первые генетические опыты Т. Моргана, которые экспериментально доказали локализацию-генов в хромосомах, а также рассмотреть построение генетических и цитологических карт и их значение.
Решите задачи 69 ... 79, приведенные в учебнике, и 84 ... 91, приведенные в методических указаниях, и внимательно их проанализируйте.
ЛИТЕРАТУРА: 1, с. 89 ... 113; 3, с. 101 ... 169.
Вопросы для самопроверки
1. Хромосомный механизм определения пола. -
2. Наследование признаков, сцепленных с полом.
3. Наследование при нерасхождении половых хромосом.
4. Основные положения хромосомной теории наследственности.
5. Сцепленное наследование, число групп сцепления.
6. Кроосинговер, механизм перекреста и обмена гомологичных хромосом в профазе редукционного деления.
7. Линейное расположение генов и частота кроссинговера.
8. Роль кроссинговера.
9. Расщепление гибридов в F 2 при .независимом комбинировании генов, полном и неполном сцеплении,
10. Генетические и цитологические карты хромосом/ их значение.
Тема 4. НЕХРОМОСОМНАЯ НАСЛЕДСТВЕННОСТЬ
Цель и задачи темы. Раскрыть роль в наследственности ■организмов, кроме ядерных, также и других генов, находящихся в цитоплазме — илазмагенов, входящих в самовоспроизводящиеся органоиды цитоплазмы (пластиды и митохондрии) и показать молекулярные основы цитоплазматической наследственности, особенности наследования признаков, контролируемых плазмогенами, пластидную наследственность, цитоплазматическую мужскую стерильность и ее использование.
Проработав тему, необходимо уметь:
— определить, по какой — отцовской или материнской — линии будет наследоваться окраска пластид;
— определить соотношение фертильных и стерильных растений в разных скрещиваниях;
— определить генетическую систему растения отцовской формы в разных скрещиваниях;
— определить, какая генетическая система отцовской линии будет полностью восстанавливать фертильность по пыльце линии;
— определить, какая генетическая система фертильной отцовской линии будет закреплять стерильность линии, скрещиваемой с этой отцовской линией.
Как уже отмечалось, основной наследственный материал эукариот сосредоточен в генах хромосом, т. е. в ядре клетки. Поэтому гены эукариот, локализованные в хромосомах, называют ядерными генами. Но чаще применяют термин «ген» без прилагательного и подразумевается при этом ядерный ген.
Однако еще в 1908—1909 гг. Корренгом и Бауром было установлено, а в наше время подтверждено, что небольшая часть наследственного материала содержится и в цитоплазме. Гены, расположенные в цитоплазме, называют цитоплазма-тическими генами, или плазмагенами.
В роли генетических элементов цитоплазмы выступают еебольшие, подобно бактериальным, кольцевые хромосомы,
Которые находятся в пластидах и митохондриях, а также генетический аппарат внутриклеточных паразитов и симбионтов (простейших, бактерий, вирусов).
Поскольку у высших растений (и животных) цитоплазмя зиготы происходит в основном от женской гаметы (яйцеклетки), то и обусловленные плазмагенами признаки передаются наиболее часто по материнской линии, как у кукурузы и других растений. Есть и второй тип передачи плазмагенов — от обоих родителей, но с необычным, неменделевским расщеплением потомства (у энотеры).
Плазмагенами определяется пестролистность кукурузы и других растений, цитоплазматическая мужская стерильность (ЦМС).
Следует обратить внимание на взаимосвязь генов хромосом и плазмагенов в формировании ряда признаков, в частности в наследовании ЦМС, и практическое использование. ЦМС в селекции.
ЛИТЕРАТУРА: 1, с. 114 ... 124, 289 .. . 294; 2, с. 144 ... 147..
Вопросы для самопроверки
1. Плазмагены, их материальная природа.
2. Роль пластид и митохондрий в наследственности.
3. Особенности наследования признаков, контролируемых плазмагенами.
4. Цитоплазматическая мужская стерильность и ее использование для получения гибридных семян.
Тема 5. МОЛЕКУЛЯРНЫЕ ОСНОВЫ ГЕНЕТИКИ
Цель и задачи темы. По данным, полученным исследователями, показать, что ДНК является основным материальным носителем наследственной информации, раскрыть структуру и функции нуклеиновых кислот, репликацию ДНК, транскрипцию и трансляцию, генетический код, синтез белка в. клетке и его регуляцию, современное представление о гене,, задачи и проблемы генетической инженерии.
Проработав тему, надо уметь:
— описать строение нуклеотида;
— описать строение молекулы ДНК и объяснить, почему число остатков аденина в молекуле ДНК равно числу остатков тимина, а число остатков гуанина равно числу остатков; цитозина;
— построить последовательность нуклеотидов в одной из цепочек молекулы ДНК по предлагаемой (Последовательности нуклеотидов второй цепочки этой же молекулы;
— описать генетический код и объяснить, почему он должен быть триплетным;
— определить комплементарную нуклеотидную последовательность матричной РНК, исходя из данных кодирующей цени ДНК и таблицы кодонов, а также указать какие кодоны из этой последовательности 'будут участвовать в синтезе пептида и какая аминокислотная последовательность образуется т результате трансляции;
— охарактеризовать роль ДНК, транспортной РНК, рибосом и аминокислот в процессе белкового синтеза;
— перечислить отдельные этапы белкового синтеза на уровне рибосом;
— рассказать, как регулируется трансляция ДНК в РНК _у прокариот;
— рассказать, какие задачи и проблемы стоят перед генетической инженерией.
При изучении темы 1 вы уже познакомились с составом и строением хромосом. Хромосомы в основном состоят из нук-леапротеидов, преимущественно из дезоксирибонуклеопротеи-дов (ДНП). В состав ДНП входят дезоксирибонуклеиновые кислоты (ДНК) и белки.
Молекула ДНК, по Уотсону и Крику, полинуклеотид и > состоит из двух длинных цепей нуклеотидов, которые образуют структуру, напоминающую винтовую лестницу. Молекула с такой структурой является двойной спиралью. В состав нуклеотида каждой из цепей входит остаток фосфорной кислоты (фосфат), пентозный сахар — дезоксирибоза и азотистые основания, представленные производными пуринов — адени-,'Ном и гуанином (А и Г) и пиримидинов — тимином и цито-зином (Т и Ц).
Содержание пуринов в ДНК всегда равно содержанию пиримидинов. В пределах пуриновой группы содержание аде-нина и гуанина может быть различно, так же, как в пределах пиримидинов содержание тимина и цитозина.
Пуриновые и пирймидиновые основания одной цепочки имеют водородные связи с пир.имидиновыми и пур.иновыми ►основаниями комплементарной цепочки. Комплементарность шх заключается в том, что в цепи всегда против тимина рас-шоложен аденин, а против гуанина.— цитозин.
ДНК является носителем наследственной информации,, которая записана в молекуле ДНК трехбуквенным кодом; на основе четырех типов мононуклеотидов.
'Смысловой единицей кода является триплет, состоящий из трех расположенных последовательно в цепи ДНК нуклео--тид.ов. Генетический код вступает в силу в процессе трансляции — синтеза белка. Триплет кодирует одну аминокислоту и называется ко доном.
Информационное значение имеет (порядок чередования нук-леотидов, содержащих разные азотистые основания. Учитывая, что в цепочке ДНК в среднем около 10 000 нуклеотидных единиц, то число молекул, различающихся по. порядку чередования оснований, выразится величиной 410000. На основе такого большого многообразия может быть записан практически любой объем информации.
Рассматривая реализацию генетической информации вч процессе биосинтеза белков, следует выяснить сущность транскрипции и трансляции. Генетическая информация о белкаж сосредоточена в хромосомах ядра клетки, а синтез белков осуществляется в рибосомах цитоплазмы. Поэтому информация с ядра должна поступить (в цитоплазму. Для этого ДНК. с помощью ферментов строит на участках своей нити молекулы информационной РНК (процесс транскрипции). Затем молекулы и-РНК переходят в цитоплазму и передают код гена на синтез белка (процесс трансляции). Следует изучить схему белкового синтеза и характер регуляции биосинтеза белков в клетке.
Следует рассмотреть подробно 'современное представление о гене, о реализации генотипа в онтогенезе, вопросы генетической инженерии.
Решите и проанализируйте задачи 85 ... 93, приведенные в учебнике и 51 ... 60, приведенные в методических указаниях.
ЛИТЕРАТУРА: 1, с. 125 ... 173; 3, с. 260 ... 356, 520 . . . 540.
Вопросы для самопроверки
1. ДНК — материальный носитель наследственной информации.
2. Синтез белка в клетке.
3. Транскрипция и трансляция.
4. Генетический код.
5. Регуляция белкового синтеза.
6. Строение и функции гена.
7. Химический и ферментативный синтез генов. Выделение
генов.
8. Современное представление о гене.
.9. Генетическая инженерия.
Тема 6. МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ
"Цель и задачи темы. Изучить основные типы мутационной изменчивости, их классификацию, индуцированный мутагенез, физические и химические мутагены, мутагены среды, проблему прогнозирования и предотвращения возможных генетических последствий, использование индуцированного мутагенеза в селекции.
Проработав тему, надо уметь изложить своими словами -шее перечисленные положения темы.
Модификационную (ненаследственную) изменчивость, а из •наследственной — комбинационную, или гибридную, мы рассмотрели в предыдущих темах.
Было показано, что при модификационной изменчивости под воздействием условий окружающей среды изменяется ■фенотип (признаки и свойства) организма, а изменений генотипа не происходит. То есть модификационная изменчивость— это норма реакции генотипа данного организма на изменение условий среды.
При гибридной (комбинационной) изменчивости изменяется (генотип потомков в результате разного сочетания и взаимодействия генов родительских форм, но новых генов при этом не образуется.
Мутационная изменчивость — это скачкообразное изменение генотипа, вызванное действиями факторов среды. Факторы, которые вызывают мутации, называют мутагенами.
По характеру изменения генетического аппарата мутации подразделяют на несколько типов: геномные мутации — изменение числа хромосом, сегментные мутации—перестройки хромосом и генные или точечные мутации.
По своей природе мутагенные факторы делят на физичес-.кие и химические. Следует хорошо усвоить характер действия -этих мутагенов на организмы, уметь при необходимости квалифицированно их использовать, не допускать отрицательного их воздействия. Химические вещества с мутагенным эффектом необходимо исключить из состава пестицидов, гербцидов, удобрений и других (Средств химизации в сельском и других отраслях народного хозяйства.
Необходимо строго соблюдать правила охраны труда при работе с химическими веществами и правила санитарной гигиены (В быту.
Среди множества разнообразных наследственных изменений (Виды и роды, генетически близкие, характеризуются сходными рядами наследственной изменчивости ,с такой правильностью, что зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Этот закон гомологических рядов в наследственной изменчивости сформулировал Н. И. Вавилов. Закон гомологических рядов позволяет селекционерам ориентироваться в создании нужных каких-то форм вида, если такие формы имеются у родственного вида. Онтогенетическая и корреляционная изменчивости рассматриваются в теме № 10.
ЛИТЕРАТУРА: 1, с. 174 .. . 227; 2, с. 89 .. . 98; 3, с. 192 .. . 257. '
Вопросы для самопроверки
1. Типы изменчивости.
2. Ненаследственная модификационная изменчивость. Норма реакции.
3. Статистические методы изучения изменчивости.
4. Наследственная изменчивость: комбинационная (гиб
ридная) и мутационная.
5. Мутации, их классификация.
6. Физические и химические мутагенные факторы, особенности их действия на наследственность.
7. Индуцированный мутагенез.
8. Закон гомологичных рядов Н. И. Вавилова.
9. Использование мутагенеза в селекции и производстве.
10. Предотвращение мутагенного загрязнения среды и со
хранение генофонда человека, животных и растений.
Тема 7. ПОЛИПЛОИДИЯ И ДРУГИЕ ИЗМЕНЕНИЯ
ЧИСЛА ХРОМОСОМ
Цель и задачи темы. Раскрыть понятие о .полиплоидии, показать морфобиологические особенности полиплоидов, их классификацию и роль в эволюции и селекции, .механизм изменения числа хромосом, использование колхицина, особенности мейоза у автополиплоидов, характер расщепления тет-раплоидных форм, триплоидию, типы аллополиплоидов, ржано-пшеничные амфидиплоиды, механизмы (возникновения и типы
анеуплоидов, особенности и классификация гаплоидов, роль разных видов полиплоидов в селекции. Проработав тему, необходимо уметь:
— определить плоидность растений;
— определить, какие типы гамет образуют разные полиплоиды; .
— обозначить, пользуясь знаками X и п, диплоидное число хромосом разных видов плоидности разных растений.
Как известно, все организмы одного биологического вида, несмотря на их возможные различия, имеют одинаковые геномы — характерную видовую совокупность постоянного числа хромосом гаплоидного набора.
В то же время могут происходить геномные мутации, связанные с изменением (увеличением или уменьшением) числа хромосом (плоидией) клеточного ядра. Под полиплоидией в широком смысле слова понимают изменение числа хромосом вообще.
Увеличение числа хромосом за счет генома одного биологического вида называют автополиплоидией (самополиплоидией), а если изменение числа хромосом происходит при межвидовой гибридизации в результате кратной суммы основных чисел геномов скрещиваемых видов, то ее называют аллопо-липлоидией (от греческого аллос— другой).
При некратном изменении числа хромосом в ядре по отношению к основному их числу в геноме называют анеуплоидией.
Увеличение числа хромосом в ядре приводит к увеличению объема клеток и до оптимального уровня плоидности увеличивается размер растений и его органов. Если число хромосом выше оптимального уровня плоидности, то такие растения развиваются хуже.
Лучше реагируют на удвоение числа хромосом виды, у которых небольшое число хромосом (например рожь), а для некоторых видов (сахарная свекла, арбуз) оптимальным уровнем плоидности является триплоидный уровень. Но триплоиды стерильны, чем затрудняется семеноводство.
При скрещивании двух разных видов или родов обычно получается бесплодное потомство, поскольку у них вследствие неродственных геномов конъюгация хромосом нормально проходить не может, и образуются нежизнеспособные гаметы. Следует обратить внимание на работы Г. Д. Карпеченко по отдельной гибридизации и механизм восстановления плодовитости межродовых гибридов путем удвоения (полиплоиды-
хзации) их хромосомного комплекса и привести достигнутые ^результаты по разным культурам.
1 Анеуплоидные организмы происходят от гамет с измененным количеством хромосом. Эти изменения .возникают у всех нормальных диплоидных организмов вследствие случайных нарушений мейоза. Анеуплоидные гаметы чаще оказываются нежизнеспособными, но некоторые их типы функционируют и дают начало анеуплоидным зиготам. Некоторые из таких зигот развиваются в анеуплоидные взрослые особи. У таких анеуплоидных организмов могут быть: тетрасомики, у которых одна из хромосом генома представлена четыре раза и организм имеет на две хромосомы больше по сравнению с диплоидом — 2n1*+2, трисомики—• 2n1* + l, моносомики — 2n1*—1, нуллисомики — 2n1*—2, тогда как нормальный дисо-мик имеет 2n.
Следует заметить, что лишняя хромосома в какой-то гомологичной паре кариотипа менее отрицательно сказывается на организм, чем ее недостаток. Так, у пшеницы наиболее жизнеспособны тетрасомики, затем в порядке понижения жизнеспособности следуют трисомики, моносомики и нуллисоми-ки. Растения, являющиеся нуллисомиками, выживают в крайне редких случаях. Все анеусомики характеризуются частичной или полной стерильностью.
Считается, что у человека возникает примерно 6% анеуплоидных зигот, (гамет, следовательно, больше) и небольшая их доля развивается во взрослые особи, обладающие рядом физических и психических недостатков. Это болезнь Дауна у человека — трисомия по 21 паре хромосом, синдром Шере-шевского — Тернера у женщин — моносомия по половой х-хромосоме и синдром Клайнфельтера у мужчин — трисомия по х-хромосоме и другие.
Гаплоиды — организмы с гаплоидным, как и в гаметах, числом хромосом —■ п. Гаплоиды развиваются из одной клетки стенотипом гаметы, минуя оплодотворение: из яйцеклетки синергиды, антиподы, пыльцевого зерна или из клеток пыльника. Гаплоиды, как правило, имеют пониженную жизнеспособность и полную или почти полную стерильность.
В генетике и селекции растений гаплоидие придается очень большое значение, поскольку этим путем можно быстро (за 2—3 года) получить максимально гомозиготные диплоидные линии (удвоить число хромосом у гаплоидных растений). При
* I — номер гомологичной пары хромосом соответствующего кариотипа.
использовании инбридинга для этого требуется не менее 5—у 6 лет и более. Но даже при длительном инбридинге не уда/ ется добиться полной гомозиготности, и та или иная степень гетерозиготности сохраняется.
Гаплоидия применяется и при отдаленной гибридизации, ее используют и для отбора рецессивных мутаций сразу после воздействия мутагенами.
Для закрепления материала решите задачи 180...-185, приведенные в задачнике [4].
ЛИТЕРАТУРА: 1, с. 228 ... 256; 2, с. 98 . .. 124; 3, с. 193 .. . 198, 344, 444... 499; 4, с. 29 ... 30.
Вопросы для самопроверки
1. Понятие о полиплоидии и полиплоидных рядах.
2. Автополиплоиды, методы их получения, использование в селекции.
3. Аллополиплоиды и их роль в селекции и растениеводстве.
4. Значение работ Г. Д. Карпеченко по отдаленной гибридизации и восстановлению плодовитости межродовых гибридов.
5. Анеуплоиды и их использование в генетике и селекции.
6. Заболевания человека, вызванные анеуплоидией.
7. Гаплоидия, методы получения, перспективы их использования в генетике, селекции, семеноводстве.
Тема 8 ОТДАЛЕННАЯ ГИБРИДИЗАЦИЯ
Цель и задачи темы. Раскрыть понятие об отдаленной гибридизации, причины нескрещиваемости видов и родов, бесплодие отдаленных гибридов и способы их преодоления, особенности формообразования в потомстве отдаленных гибридов, использование отдаленной гибридизации в селекции.
Проработав тему, необходимо уметь:
— дать определение понятию отдаленной гибридизации;
— назвать причины нескрещиваемоети видов и родов;
— объяснить причины бесплодия и указать особенности формирования отдаленных гибридов;
— указать способы преодоления нескрещиваемости видов и родов и бесплодия отдаленных гибридов;
— привести примеры использования отдаленной гибридизации в селекции растений.
У биологического вида его генетическая система представлена видоспецифическими генетическими структурами, которые на основе нормальных процессов мейоза и оплодотворения устойчиво воспроизводятся в поколениях « надежно защищены от проникновения в них структурных элементов других видов.
Генетическая обособленность системы особей одного вида от других обеспечивается наличием барьеров, которые исключают или сильно ограничивают обмен генетической информацией между разными видами.
Основной причиной нескрещиваемости представителей разных биологических видов является несбалансированность их 'кариотипов и генотипически обусловленная физиологическая несовместимость. В разных случаях она проявляется специфически и выражается в остановке процесса оплодотворения на том или ином этапе.
В то же время отдаленная гибридизация — межвидовая, межродовая, а также скрещивание сильно отличающихся форм одного биологического вида представляет не только теоретическое, но и практическое значение, поскольку можно соединить в гибриде ценные свойства разных видов, родов и других скрещиваемых сильно отличающихся форм.
Первое поколение межвидовых и межродовых гибридов примерно одинаково похожи на обоих родителей или немного уклоняются в сторону от них и, как правило, характеризуется или полным бесплодием, или в разной степени пониженной, по сравнению с нормальной, плодовитостью.
В последнем случае фертильность при размножении гибридов обычно постепенно повышается, но межродовые гибриды (расщепляются с возвратом к исходным родительским видам. Рекомендации разных видовых свойств происходят в редких случаях.
Нескрещиваемость разных биологических видов, да и родов, можно в определенной степени преодолеть разными 'способами. Для этого необходимо изучить особенности мейоза и специфику формообразовательного процесса у отдаленных гибридов.
Стерильность, вызванную различными нарушениями мейоза, можно преодолеть удвоением числа хромосом — полипло-идизацией и деполиплоидизащией исходных форм, сбалансировав этим путем их кариотипы.
В какой-то степени способствует преодолению нескрещиваемости <
|
из
5.00
|
Обсуждение в статье: Тема 2. ЗАКОНОМЕРНОСТИ НАСЛЕДОВАНИЯ ПРИЗНАКОВ |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы