 |
Главная |


ПРИНЦИПЫ ФУНКЦИОНИРОВАНИЯ БИОЛОГИЧЕСКИХ СИСТЕМ
|
из
5.00
|
Структурно-функциональные принципы. Экспериментальные исследования биологических систем обычно сопровождаются теоретическими обобщениями. При этом применение математических методов анализа способствует более строгому формулированию общих положений.
Ранний период внедрения математических методов в биологию относится ко второй половине прошлого века, когда начала разрабатываться вариационная статистика, которую «...Пирсон и его продолжатели специально развивали и расширяли, чтобы сделать ее алгоритмы как можно более пригодными к оценкам, сравнениям и количественным характеристикам биологических явлений»
[36].
В начале XX в. П. П. Лазарев, В. Нернст иДж.Леб,
исследуя раздражения нервной ткани, установили количественные закономерности и вывели формулы, связывающие величину возбуждения с силой электрического раздражения. Несколько ранее, в 1901 г., Ж. Вейс пред дожил использовать для этой цели формулу гиперболы. Таким образом, в области нейрофизиологии давно ощущалось влияние математизации, что сказалось как на проведении самих экспериментов, так и на формулировании результатов. Если результаты и не облекались в строгую математическую форму, то формулировались они достаточно логично.
Процесс обобщения результатов и формулирования общих положений, связанных с функционированием биосистем, усилился с развитием кибернетики и внедрением в биологические исследования методов моделирования. Общие принципы, выдвинутые кибернетиками при изучении работы технических систем, оказались вполне применимыми к биологическим.
Рассмотрим группу принципов, связанных со структурой и функцией биосистем, между которыми существует тесное единство. В каждый момент времени функция формируется на структуре, однако необходимость выполнять определенную функцию в течение некоторого времени неизбежно приводит к формированию новой структуры.
Одним из общих принципов, связывающих структуру и функцию системы, является принцип наипростейшей конструкции, сформулированный в 1943 г. американским биофизиком Н. Рашевским. Одни и те же функции при одинаковой их интенсивности, вообще говоря, могут выполняться несколькими различными структурами. Согласно принципу Рашевского [114] «...та конкретная структура или конструкция, которую мы действительно находим в природе, является простейшей из возможных структур или конструкций, способных выполнять данную функцию или группу функций». В 1954 г. американский ученый Д. Кон усилил формулировку этого принципа и предложил принцип оптимальной конструкции, по которому органическая структура, необходимая для выполнения данной функции, должна быть оптимальной в отношении нужного количества материала и необходимых затрат энергии. Дальнейшее изучение структурных особенностей биосистемы привело Н. Рашевского в 1961 г. к принципу адекватной конструкции организма [114]: «Конструкция должна быть адекватной заданной функции при заданных изменяющихся условиях среды».
Сформулированные выше принципы допускают, очевидно, не единственное решение проблемы формы и струк туры. Но это является не недостатком, а преимуществом. В органическом мире действительно существуют организмы и органы, которые выполняют в основном одинаковые функции, но тем не менее различны по форме. Однако нельзя утверждать, что системы, выполняющие разные функции, всегда различны по своей структуре.
Дополнением к теории адекватной конструкции Рашевского является принцип эволюционной компенсации силы тяжести [28]. Применительно к рецепторному аппарату вестибулярного анализатора он формулируется следующим образом: в условиях нормальной весомости сила тяжести компенсируется внутренними упругими силами конструкции рецепторов. В периферической части вестибулярного анализатора в процессе эволюции этот принцип используется для увеличения чувствительности рецепторных окончаний специфических нервных клеток.
Обобщением взаимоотношений структуры и функции является принцип структурно-функционального единства. В любой биологической системе структура и функция представляют единое целое, причем функциональный эффект биосистемы достигается за счет ее внутреннего структурирования. Единство структуры и функции биосистемы формируется в процессе взаимодействия со средой и является проявлением адекватности биосистемы среде. В самой биосистеме структура и функция взаимоадекватны.
Принцип структурно-функционального единства применительно к нервной клетке был сформулирован в 1973 г. [75]: пространственно-временная функциональная неоднородность нервной клетки обеспечивается взаимодействием пространственно распределенных структурно неоднородных синаптических образований со структурно однородной мембраной клетки.
Этот принцип справедлив для биосистем любого иерархического уровня. Чтобы его установить, необходимо детально изучить структурирование биосистемы на подсистемы и проанализировать формирование функции каждой подсистемы и биосистемы в целом на основе взаимодействия между подсистемами.
Свойство иерархичности биосистем позволило сформулировать принцип этажности [61. В любой сложной системе этажи переработки информации взаимосвязаны и оказывают влияние один на другой. На определенном этапе эволюции биосистем начали возникать этажные структуры. Живые системы усложнялись сначала за счет длины цепей элементов, затем их соединения становились параллельными, далее происходила надстройка этажей.
Для клетки этажи — это атомы (если отбросить элементарные частицы), простые и сложные молекулы, структуры из молекул и, наконец, целые клетки. На каждом этаже происходит обмен энергией и вещественными частицами. В процессе развития каждой регулирующей системы организма также формируется сложная структура иерархических этажей с вертикальными связями. Одновременно закладываются горизонтальные связи между соответственными этажами близких регулирующих систем. Для организма прибавляются новые этажи — ткани, органы, системы органов, целостный организм.
Затем формируются иерархические уровни систем, объединяющие несколько первичных систем. На каждом уровне сначала образуются параллельные, затем этажные структуры. Таким образом, сложные отношения, в которых находятся между собой биологические системы организма, носят иерархический характер. Степень независимости одной системы от другой, более крупной, определяется ее жизнеспособностью при отключении от нее энергетических и информационных воздействий со стороны других подобных систем. С понятием сложных отношений связана степень упорядоченности системы или степень непротиворечивости деятельности ее подсистем и элементов, т. е. то, насколько частные функции не мешают и не противодействуют друг другу.
Повышение степени упорядоченности увеличивает устойчивость системы, но понижает способность ее к эволюции.
Принципы динамического функционирования биосистем. Одна из первых попыток формирования принципа работы биосистемы принадлежит П. Мопертюи (XVIII в.). Его принцип наименьшего действия гласит: когда в природе происходит некоторое изменение, количество действия, необходимое для этого изменения, является наименьшим возможным. По иронии судьбы принцип Мопертюи, высказанный по отношению к живой природе, к биосистемам, получил строгую трактовку уже безотносительно к биологии, и теперь широко известны егоприложения к теории синтеза технических систем автоматического управления.
Начало анализа сохранительных свойств биосистем связывается с обобщением принципа самосохранения Ле-Шателье: если на систему, находящуюся в устойчивом равновесии, подействовать извне, изменяя какое-нибудь из условий, определяющих положение равновесия, то равновесие смещается в том направлении, при котором эффект произведенного воздействия уменьшается.
Этот принцип был сформулирован французским химиком А. Ле-Шателье в 1887 г. для термодинамических систем. После того как немецкий физик К. Браун показал, что это положение является следствием второго начала термодинамики, его иногда стали называть принципом Ле-Шателье—Брауна.
Аристотель учил, что каждому элементу отвечает свое естественное место во Вселенной, к которому тела стремятся по природе с тем, чтобы тем достигнуть покоя. После Ньютона, однако, стало ясно, что такого естественного места не существует: все тела, если на них действуют силы, сохраняют свое состояние, а изменение состояния, каким бы оно ни было, осуществляется внешними силами. Тела же сопротивляются любому насильственному изменению своего состояния.
Б. Спиноза сформулировал свою концепцию самосохранения так: «Никакая вещь не имеет в себе ничего, через' что она могла бы уничтожиться; наоборот, она противодействует всему тому, что может уничтожить ее существование. Следовательно, насколько возможно и насколько это от нее зависит, она стремится пребывать в своем существовании» [125].
Принцип Ле-Шателье — Брауна, таким образом, можно трактовать как частное применение общего принципа самосохранения к термодинамическим системам.
В сходных терминах было сформулировано У. Кен-ноном и понятие самосохранения живых систем: «В открытой системе, такой как наши организмы, состоящие из нестабильного материала и подверженные непрерывному воздействию возмущений, само постоянство служит доказательством существования агентов, действующих или готовых к действию, чтобы поддержать это постоянство.
Если состояние остается устойчивым, то это происходит потому, что любая тенденция к его изменению авто магически вызывает увеличение эффективности фактора или факторов, противодействующих этому изменению» [154].
Важность принципов самосохранения для понимания процессов в живых системах очевидна. Эти принципы в различных сферах исследования используются в различных формах. Так, часто принципы сохранения принимают гомеостатический характер, в той или иной мере близкий принципу Ле-Шателье [33, 154]. В иных случаях связь концепций с принципом Ле-Шателье отрицается [38].
Большое значение для развития представлений об общих принципах работы биосистем имеет сформулированный И. М. Гельфандом и М. Л. Цетлиным принцип наименьшего взаимодействия. Согласно этому принципу, систему можно назвать целесообразно работающей в некоторой внешней среде, если система стремится минимизировать взаимодействие со средой [142]. При этом мерой взаимодействия организма со средой может служить отклонение параметров внутренней среды организма от оптимальных значений. Этот принцип был ими выдвинут при изучении механизмов управления движениями и взаимодействием иерархических уровней нервной системы. Суть его состоит в том, что если рассматривать сложную систему управления как совокупность подсистем, составляющих несколько уровней, то каждая такая подсистема стремится уменьшить свое взаимодействие с внешней средой. На каждом уровне внешние воздействия включают идущую с нижних уровней афферентацию, а взаимодействие высших уровней определяет организацию взаимодействия низших уровней. Модель, соответствующая этому принципу, определяет достижение цели в меняющейся среде с помощью методов поиска экстремума нестационарной функции многих переменных.
В развитие теоретической биологии большой вклад внес советский биофизик Э. Бауэр, изучивший химические и физические процессы, протекающие в организме на клеточном уровне. Свои выводы он изложил в виде очень оригинальных принципов, облеченных в строгую математическую форму, в книге «Теоретическая биология» (1935 г.). Э. Бауэр подчеркивал, что на уровне законов химии и физики в организме не наблюдается равновесия, напротив, организму присуще устойчивое неравновесие. Рассматривая процессы обмена между биосистемой и средой по веществу и энергии, Э. Бауэр [32] формулирует некоторые принципы, общие для живых систем:
всем живым существам свойственно прежде всего самопроизвольное изменение своего состояния, т. е. изменение состояния, которое не вызвано внешними причинами, лежащими вне живого организма;
при изменении внешних условий существо не просто противодействует внешней силе (инерции и трению по принципу Даламбера), а в результате противодействия изменяет состояние среды;
работа живых систем при всякой окружающей среде направлена против равновесия, которое должно было бы наступить в данной окружающей среде при данном первоначальном состоянии системы.
Обобщением этих трех принципов является сформулированный Э. Бауэром всеобщий закон биологии: все и только живые системы никогда не бывают в равновесии и исполняют за счет своей свободной энергии постоянно работу против равновесия, требуемого законами физики и химии при существующих внешних условиях. Этому закону он придал следующую математическую трактовку:

где F — свободная энергия системы при наступившем равновесии; х— разность факторов работы (разность потенциалов, концентраций); Dх— изменения, вызванные этими факторами; Dl — время, в течение которого произошли эти изменения. Здесь штрих относится к неживой системе, а разность в правой части соответствует работе живой системы против выравнивания.
Практически всему человечеству известны работы И. П. Павлова по условным рефлексам. Здесь мы лишь обсудим соотношение взглядов И. П. Павлова с понятиями кибернетики. Практически все работы И. П. Павлова и его школы относятся к исследованию тех или иных форм поведения, к исследованию условий, в которых под влиянием изменившейся окружающей среды разрушается старый и формируется новый стереотип поведения. Известно также, что в кибернетике принято рассматривать систему и ее среду и объяснять изменение поведениясистемы изменением среды. С этой точки зрения исследование формирования простых, а затем и сложных инструментальных рефлексов у животных под действием безусловных и условных раздражителей есть исследование кибернетических (живых) систем. И результаты, полученные в этом отношении акад. И. П. Павловым, трудно переоценить. Он не использовал математические методы при интерпретации своих экспериментов, но ему удалось сформулировать ряд весьма важных принципов, которые до сих пор не утратили своего значения.
При исследовании взаимодействия организма со средой И. П. Павлов обосновал логическую структурную модель условных рефлексов. В эксперименте он создал специальные комбинации входных воздействий на организм, четко регистрировал реакцию организма не только по общему поведению, но и по качественным значениям физиологических показателей.
Результатом такого структурного подхода явилось формулирование И. П. Павловым в конце 20-х и в начале 30-х годов принципа уравновешивания системы со средой, который обобщал три важных положения [104]:
мозг регулирует процессы, протекающие внутри организма,— принцип нервизма;
мозг устанавливает и регулирует взаимосвязь между организмом и внешней средой на базе безусловных и условных рефлексов — принцип условных и безусловных рефлексов;
кора головного мозга осуществляет функцию образования и сохранения динамического равновесия между внутренней средой организма и окружающей организм средой — принцип динамического уравновешивания внешней и внутренней среды.
Если обратиться к истокам кибернетики, нельзя не отдать должное исследованиям И. П. Павлова. И принцип нервизма, и принцип безусловных и условных рефлексов, и принцип уравновешивания внешней и внутренней среды привели к выводу о том, что биологическая система, активно перестраивая свое поведение, стремится к уравновешиванию со средой. Это дает ключ к пониманию не только динамического приспособления к изменившимся условиям среды в онтогенезе, при обучении, но и в эволюции видов, к филогенезу.
Акад. И. П. Павлов является также основателем школы, использующей метод логического структурирования биологической модели при исследовании биосистем различного иерархического уровня.
Принцип функциональной системы. В 30-е годы идеи, связанные с принципом и теорией функциональной системы, развивал акад. П. К. Анохин. Он придал законченный вид идее рефлекторной дуги И. П. Павлова, характерный для систем с обратной связью. Им был выдвинут и разработан тезис об обратной афферентации— своеобразном замыкании обратной связи организма через окружающую среду — и подчеркнут сложный характер обработки информации в цепи обратной афферентации. Фундаментальным является и положение П. К. Анохина о роли результата как фактора, образующего функциональную систему. Как он справедливо отмечает, все определения систем, бытующие даже сейчас в кибернетике и общей теории связи, являются неполными вследствие отсутствия связи работы системы с требуемым конечным результатом. Если И. П. Павлов, выдвинув принцип динамического уравновешивания организма со средой, указал на конечную естественную цель, достигаемую организмом, обитающим в данной среде и приспосабливающимся к ней, то теория функциональной системы П. К. Анохина раскрывает, как организм может достигнуть этой конечной цели, какие механизмы должны для этого действовать. Любопытно отметить, что идеи П. К. Анохина, связанные с анализом биосистем, т. е. наиболее сложных систем, опережают идеи, возникающие в кибернетике на основе анализа и усложнения технических систем, в частности необходимость замыкания обратной связи обоснована им еще в 1935 г. Обратная аф-ферентация являет собой пример наиболее сложной обработки информации, для которой в технических системах нынче используются вычислительные машины;
выдвинутое им положение об акцепторе действия предвосхитило идеи оптимального и критериального управления.
Рассмотрим основные положения общей теории функциональных систем организма. Различные этапы формирования системы фактически подчинены решению следующих вопросов: какой результат должен быть получен; когда именно должен быть получен результат;
какими механизмами должен быть получен результат;
как система убеждается в достаточности полученного результата. П. К. Анохин придает результату возможность организовать распределение возбуждений в системе в соответствующем направлении. Таким образом, все формирование системы подчинено получению определенного полезного результата; недостаточный результат может целиком реорганизовать систему и сформировать новую, с более совершенным взаимодействием компонентов, дающим достаточный результат.
П. К. Анохин дает следующее определение понятия системы. [17]: «...системой можно назвать только такой комплекс избирательно вовлеченных компонентов, у которых взаимодействие и взаимоотношение приобретают характер взаимосодействия компонентов на получение фиксированного полезного результата».
В свою очередь результат благодаря обратной аф-ферентации имеет возможность реорганизовать систему, создавая такую форму взаимодействия между ее компонентами, которая является наиболее благоприятной для получения именно запрограммированного результата. Таким образом, результат рассматривается как неотъемлемый и решающий компонент системы, как инструмент, создающий упорядоченное взаимодействие между всеми другими ее компонентами.
Функциональные системы организма складываются из динамически мобилизуемых структур в масштабе целого организма. Наличие результата системы как определяющего фактора для формирования функциональной системы и наличие специфического строения структурных аппаратов, дающих возможность немедленной мобилизации объединения их в функциональную систему, говорит о том, что системы организма всегда функциональны. Это значит, что функциональный принцип выборочной мобилизации структур является доминирующим. Поэтому такая система и была названа функциональной [15].
С вопросом структурного состава функциональной системы связан и вопрос об иерархии систем. Говоря о составе функциональной системы, следует иметь в виду, что всякая данная функциональная система, взятая для исследования, неизбежно находится где-то между тончайшими молекулярными системами и наиболее высоким уровнем системной организации.
П. К. Анохин делает следующий вывод о составе иерархии: все функциональные системы, независимо от уровня своей организации и от количества составляющихих компонентов, имеют принципиально одну и ту же функциональную архитектонику, в которой результат является доминирующим фактором.
Главной чертой каждой функциональной системы является ее динамичность. Структурные образования, составляющие функциональные системы, обладают исключительно подвижной мобилизуемостью. Именно это свойство систем и дает им возможность быть пластичными, внезапно менять свою архитектонику в поисках запрограммированного полезного результата.
Применение системы как инструмента в научных исследованиях затруднено настолько, что многие исследователи не изучают внутреннее строение системы, а ограничиваются исследованием на уровне «черного ящика». Между тем вскрытие внутренних закономерностей действия системы, ее узловых механизмов позволило бы добиться главного в исследовательском процессе: удержания в руках целого, когда анализируются части этого целого [1б].
Функциональная система всегда гетерогенна. Она состоит из определенного количества узловых механизмов, каждый из которых занимает надлежащее место и является специфическим для всего процесса формирования функциональной системы. Вскрытие этих механизмов, составляющих внутреннюю архитектонику системы, приблизит исследователей к самой решающей цели системного подхода вообще — обеспечить органическое единство в исследовательском процессе системного уровня функционирования с индивидуальной характеристикой каждого дробного элемента или механизма, принимающего участие в этом функционировании.
Одним из достоинств общей теории функциональных систем является разработка модели системы с четко отработанным внутренним строением или, по выражению П. К. Анохина, внутренней архитектоникой. Такая внутренняя архитектоника, выраженная в физиологических понятиях, является непосредственным инструментом для практического применения функциональной системы в исследовательской работе.
Рассмотрим узловые специфические механизмы, представляющие собой внутреннюю архитектонику системы (рис. 5).
Афферентный синтез. Биосистема, даже простой иерархии, сама на основе внутренних процессов прини мает решение о том, какой результат нужен в данный момент ее приспособительной деятельности. Вопрос этот решается именно на стадии афферентного синтеза.
Выдвигаются четыре решающих компонента афферентного синтеза, которые должны быть подвергнуты одновременной обработке с одновременным взаимодействием на уровне отдельных нейронов: доминирующая на данный момент мотивация; обстановочная афферентация, также соответствующая данному моменту; пусковая афферентация и, наконец, память
Основным условием афферентного синтеза является

Рис. 5. Общая характеристика функциональной системы (по П. К. Анохину).
одновременная встреча всех четырех составляющих этой стадии функциональной системы. Микроэлектродный анализ, микрохимическое исследование и другие формы аналитического исследования нейрона в момент встречи на нем упомянутых выше четырех возбуждений показали, что этот процесс поддерживается и облегчается рядом динамических процессов нервной системы. Это прежде всего выходящая активация, сопутствующая афферентному синтезу и предшествующая принятию решения [88, 141]. Сюда же относятся процесс корково-подкорковой реверберации [147] и процесс центробежного повышения возбудимости вовлеченных в афферентный синтез рецепторов.
Афферентный синтез, приводящий организм к решению вопроса, какой именно результат должен быть получен в данный момент, обеспечивает постановку цели, достижению которой и будет посвящена вся дальнейшая логика системы. Принятие решения является следующим узловым механизмом функциональной системы. Афферентный синтез, подчиняясь доминирующей на данный момент мотивации, осуществляет подбор тех значений элементов системы, при которых возбуждения избирательно направляются к мышцам, совершающим нужное действие. Любое принятие решения после окончания афферентного синтеза является выбором наиболее подходящих значений элементов, которые должны составить рабочую часть системы. Эти значения дают возможность экономно осуществить именно то действие, которое должно привести к запрограммированному результату.
Принятие решения — это в высшей степени конденсированный процесс, в котором одновременно обрабатывается на основе доминирующей мотивации вся пришедшая в мозг афферентная информация, производится непрерывное сопоставление этих результатов с прошлым ответом и переводятся результаты этой обработки на афферентные пути, соответствующие распределению возбуждений для совершения нужного акта, обеспечивающего получение необходимых результатов.
Формирование акцептора результатов действия. Акцептор результатов действия является весьма сложным аппаратом. По сути он должен сформировать определенные нервные механизмы, которые позволяют не только прогнозировать признаки необходимого в данный момент" результата, но и сличать их с параметрами реального результата, информация о которых приходит к акцептору результатов действия благодаря обратной афферента-ции. Именно этот аппарат дает единственную возможность организму исправить ошибку поведения или довести несовершенные поведенческие акты до совершенных.
П. К. Анохиным с сотрудниками был доказан в ряде экспериментов — ив обычных с условными рефлексами, и с помощью тонких электрофизиологических приемов — тот факт, что аппарат акцептора результатов действия организуется непосредственно после принятия решения.
Очевидно, что существенные признаки будущего результата динамически формируются благодаря многосторонним процессам афферентного синтеза с извлечением из памяти прошлого жизненного опыта и его результата. Этот комплекс возбуждений является афферентной моделью будущего результата, а именно эта модель, яв ляясь эталоном оценки обратных афферентаций, должна направлять активность человека и животных вплоть до получения запрограммированного результата.
В последнее время были получены сведения о том,что в этот нервный комплекс, обладающий высокой степенью мультиконвергентного взаимодействия, приходит еще одно возбуждение, уже эфферентной природы. Речь идет о коллатералях пирамидного тракта, которые по многим межуточным нейронам отводят «копии» тех эфферентных посылок, которые выходят на пирамидный тракт. Эти эфферентные возбуждения конвергируют на те же межуточные нейроны сенсомоторной области, куда поступают и все те афферентные возбуждения, которые могут составить параметры реального результата.
Таким образом, момент принятия решения и начала выхода рабочих эфферентных возбуждений из мозга сопровождается формированием обширного комплекса возбуждений, состоящего из афферентных признаков будущего результата и из коллатеральной копии эфферентных возбуждений, вышедших на периферию по пирамидному тракту к рабочим аппаратам. В зависимости от интервала между постановкой цели и ее реализацией к этому же комплексу возбуждений через определенное время приходят возбуждения и от реальных параметров полученного результата. Сам процесс оценки полученного реального результата осуществляется из сличения прогнозированных параметров и параметров реально полученного результата. Именно здесь осуществляется оценка полученного результата. Оценка и результат оценки определяют дальнейшее поведение организма. Если результат соответствует прогнозу, то организм переходит к следующему этапу поведения. Если же результат не соответствует прогнозу, то в аппарате сличения происходит рассогласование, активирующее ориентировочно-исследовательскую реакцию, которая, поднимая ассоциативные возможности мозга на более высокий уровень, тем самым помогает активному подбору дополнительной информации.
Принцип функциональной системы, являющийся достаточно общим, оказался полезным при изучении биосистем различного иерархического уровня, а детальное структурирование позволяет проводить эксперименты по выявлению взаимодействия между отдельными блоками системы. Принципы самоорганизации и адекватности. Большое внимание исследователей привлекает такое фундаментальное свойство биосистем, как способность изменять свою сложность и организацию. Это качество биосистемы изучалось в самых различных аспектах. У. Р. Эшби обратил внимание на необходимость установления соответствия между сложностью биосистемы и сложностью среды. В 1958 г. он сформулировал принцип необходимого разнообразия: только разнообразие может уничтожить разнообразие. Этот принцип означает, что биологическая система может функционировать в среде, если число состояний системы равно числу состояний среды.
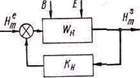
Рис. 6. Схема взаимодействия по сложности.
Развитие принципа необходимого разнообразия [25] предусматривает не для всех случаев взаимодействия системы и среды установление равновесия по числу состояний. Используя меру сложности, введенную У. Р. Эшби, можно записать функции числа состояний для биосистемы и среды:
Н s m = log ns; Hem == log ne
(1.16)
где индекс s относится к системе, индекс е — к среде.
Рассогласование между биосистемой и средой по сложности в этом случае определяется следующим образом:

где kн — коэффициент пропорциональности.
Это рассогласование является управляющим сигналом, который действует на механизм изменения сложности Wн за счет постоянного притока вещества В и энергии Е (рис. 6).
Таким образом, биосистема, используя устойчивое неравновесие, способна изменить свою структуру и функциональную сложность (число состояний). Сложность биосистемы изменяется за счет изменения размеров, числа элементов, числа связей между элементами или путем изменения параметров элементов, динамической конфигурации связей между ними. Изменение функциональной сложности может быть связано с использованием ранее не задействованных структурных элементов, с изменением порядка и разнообразия реакций биосистемы Измерение числа состояний среды может во многих случаях непосредственно производиться биосистемой. При этом возможны три случая взаимодействия системы со средой в зависимости от величины kн:
1) kн = 1 — каждому состоянию среды соответствует состояние биосистемы — прямое использование принципа необходимого разнообразия Эшби;
2) kн < 1 — каждому состоянию среды может соответствовать некоторое множество состояний биосистемы; такой случай возможен, если биосистема обладает возможностью тонкого и структурного анализа состояний среды;
3) kн > 1 — определенному набору числа состояний среды соответствует некоторый меньший набор числа состояний биосистемы; этот случай может соответствовать процессу укрупнения показателей среды по тем или иным критериям, процессу обобщения в образ.
При анализе разнообразия системы и среды нельзя исключить ни один из рассмотренных выше случаев. Более того, найденное тем или иным способом экспериментальное значение kн может указать на характер взаимодействия биосистемы со средой (анализ, синтез, взаимно-однозначное соответствие). Например, если взаимодействие биосистемы со средой проходит в условиях дефицита вещества и энергии, то в биосистеме может происходить вынужденное обобщение числа состояний среды. Динамика установления необходимого разнообразия в биосистеме может быть различной. Если необходимое разнообразие может быть достигнуто за счет мобилизации структурных и (или) функциональных резервов, то установление может осуществиться достаточно быстро. Если же для необходимого разнообразия биосистема должна заниматься перестройкой собственной структуры, то этот процесс может быть достаточно длительным. В соответствии с этим основанием для перестройки структуры является частое и длительное воздействие среды на биосистему.
Рассматривая возможности биосистемы по изменению уровня организации, У. Р. Эшби [150] отметил, что самоорганизация как процесс может означать переход от неорганизованной системы к организованной или переход от плохо организованной системы к хорошо организованной. Он предложил считать критерием самоорганизации биосистем убывание неопределенности, выражаемое условием

В 1960 г. Г. Ферстер ввел в рассмотрение оценку относительной организации и предложил считать систему самоорганизующейся, если

Усложнение простых условий взаимосвязи биосистемы и среды и учет влияния среды на сложность системы приводят к необходимости рассмотрения текущей неопределенности системы как функции распределения вероятностей состояний. В этом случае условия самоорганизации принимают вид [19]:

Рассмотренные выше условия изменения энтропии (неопределенности) биосистемы обладают общей следующей особенностью: влияние среды оценивается лишь по изменению неопределенности или уровня организации биосистемы.
Более четко принцип самоорганизации был сформулирован в 1962 г. В. М. Глушковым. При этом рассматривалось взаимодействие системы (в том числе и биосистемы)" со средой и изменение неопределенности системы как функция этого взаимодействия. Пусть Q представляют собой сигналы среды, воспринимаемые биосистемой как обучающая последовательность. Среди всех обучающих последовательностей есть последовательность Q0, при которой энтропия биосистемы максимальна. Вообще говоря, обучающую последовательность Q0 можно считать тем первоначальным контактом, который биосистема установила с данной средой. Понятно, что если биосистема способна обучаться, то каждый последующий контакт со средой может уменьшать неопределенность системы. Тогда изменение неопределенности
D HQ=HQ-HQ0 (1.21)
является величиной отрицательной.
Часть обучающих последовательностей среды может использоваться или считаться экзаменационными. Пусть биосистема многократно (k раз) может подвергатьсявыражаемое условием

В 1960 г. Г. Ферстер ввел в рассмотрение оценку относительной организации и предложил считать систему самоорганизующейся, если
Усложнение простых условий взаимосвязи биосистемы и среды и учет влияния среды на сложность системы приводят к необходимости рассмотрения текущей неопределенности системы как функции распределения вероятностей состояний. В этом случае условия самоорганизации принимают вид [19]:
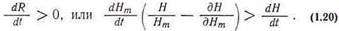
Рассмотренные выше условия изменения энтропии (неопределенности) биосистемы обладают общей следующей особенностью: влияние среды оценивается лишь по изменению неопределенности или уровня организации биосистемы.
Более <
|
из
5.00
|
Обсуждение в статье: ПРИНЦИПЫ ФУНКЦИОНИРОВАНИЯ БИОЛОГИЧЕСКИХ СИСТЕМ |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы