 |
Главная |


Синаптическая передача информации
|
из
5.00
|
Уже отмечалась важная роль мембраны в передаче информации в мозге. Мембрана представляет собой барьер для прохождения нервного импульса. Имен-
| Рис. 1.17. Структура синапса (а); фотография синапса, увеличенного под электронным микроскопом в тысячи раз (выполненная D.D. Kinkel) (б); фотография аксональных терминалей на теле аксона (в) (Kalat, 1992). |
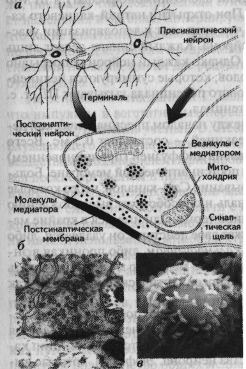 но поэтому связи между нейронами опосредуются химическими передатчиками — нейромедиаторами (mediator — посредник, англ.), выделяющимися из окончаний аксонов в области специализированных межклеточных контактов — синапсов. Синапс представляет собой мембраны двух соседних нейронов (передающего информацию и получающего ее) и пространство между ними, которое называется си-наптической щелью. Синаптическая щель — это пространство шириною около 20 нм между мембранами пре-синаптической (мембрана нейрона, находящегося перед синаптической щелью) и постсинаптической (мембрана клетки, находящейся после синаптической щели) клеток (рис. 1.17). Различают аксо-соматические синапсы, сформированные мембранами аксона и тела (сомы) нейрона, аксо-дендритные, состоящие из мембраны аксона и дендрита, и аксо-ак-сональные, при которых аксон подходит к другому аксону. Синапс между аксонами и мышечным волокном называется нейромышечной кольцевой пластинкой.
но поэтому связи между нейронами опосредуются химическими передатчиками — нейромедиаторами (mediator — посредник, англ.), выделяющимися из окончаний аксонов в области специализированных межклеточных контактов — синапсов. Синапс представляет собой мембраны двух соседних нейронов (передающего информацию и получающего ее) и пространство между ними, которое называется си-наптической щелью. Синаптическая щель — это пространство шириною около 20 нм между мембранами пре-синаптической (мембрана нейрона, находящегося перед синаптической щелью) и постсинаптической (мембрана клетки, находящейся после синаптической щели) клеток (рис. 1.17). Различают аксо-соматические синапсы, сформированные мембранами аксона и тела (сомы) нейрона, аксо-дендритные, состоящие из мембраны аксона и дендрита, и аксо-ак-сональные, при которых аксон подходит к другому аксону. Синапс между аксонами и мышечным волокном называется нейромышечной кольцевой пластинкой.
| Рис. 1.18. Временная и пространственная суммация импульсов (Kalat, 1992). |
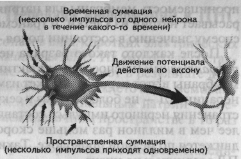 Молекулы медиатора находятся в везикулах — особых пузырьках, расположенных в аксональной терми-нали (окончании аксона). ПД, достигая аксональной терминали, становится сигналом открытия кальциевых каналов, которые вызывают синхронный эндоцитоз — координированное выделение медиатора из везикул и поступление их в синапти-ческую щель. Медиатор связывается с рецептором, находящимся на пост-синаптической мембране, который инициирует в постсинаптической клетке те или иные изменения в зависимости от вида рецептора. Медиатор, взаимодействуя с рецептором, может способствовать открытию ионных каналов (натрий-калиевых или кальциевых) или через аденилатциклазный механизм активировать внутриклеточного посредника — цАМФ (циклический аденозинмонофосфат) и цГМФ (циклический гуанозинмонофосфат). При открытии натрий-калиевого канала натрий поступает внутрь клетки, что приводит к деполяризации участка мембраны постсинаптического нейрона. Каждый синапс делает лишь незначительный вклад в этот процесс. Однако каждый нейрон непрерывно интегрирует до 1000 синаптических входов, которые суммируются нелинейно (рис. 1.18) и при достижении порогового потенциала вызывают ПД, т. е. распространяющийся вдоль аксона потенциал.
Молекулы медиатора находятся в везикулах — особых пузырьках, расположенных в аксональной терми-нали (окончании аксона). ПД, достигая аксональной терминали, становится сигналом открытия кальциевых каналов, которые вызывают синхронный эндоцитоз — координированное выделение медиатора из везикул и поступление их в синапти-ческую щель. Медиатор связывается с рецептором, находящимся на пост-синаптической мембране, который инициирует в постсинаптической клетке те или иные изменения в зависимости от вида рецептора. Медиатор, взаимодействуя с рецептором, может способствовать открытию ионных каналов (натрий-калиевых или кальциевых) или через аденилатциклазный механизм активировать внутриклеточного посредника — цАМФ (циклический аденозинмонофосфат) и цГМФ (циклический гуанозинмонофосфат). При открытии натрий-калиевого канала натрий поступает внутрь клетки, что приводит к деполяризации участка мембраны постсинаптического нейрона. Каждый синапс делает лишь незначительный вклад в этот процесс. Однако каждый нейрон непрерывно интегрирует до 1000 синаптических входов, которые суммируются нелинейно (рис. 1.18) и при достижении порогового потенциала вызывают ПД, т. е. распространяющийся вдоль аксона потенциал.
Синоптическая задержка — время между началом пресинаптической деполяризации и постсинаптической реакцией — составляет 0,5 мс. Всего лишь 1/10 часть этого времени обусловлена диффузией (проникновением) медиатора через синаптическую щель к постсинаптической мембране. Большая же часть времени тратится на открывание Са2+-каналов, через которые кальций проникает в аксонную терминаль и способствует высвобождению медиатора из везикул. Кальций в аксонной терминали имеется в крайне малых количествах. После того как он окажет свое действие, он удаляется, либо связываясь со специальным белком — кальмодулином, либо проникая в эн-доплазматический ретикулум. Освобожденный медиатор может действовать как на ауторецепторы (расположенные на пресинаптической мембране), так и на постсинаптические рецепторы.
Согласно принципу английского ученого Г. Дейла, постулирующего метаболическое единство нейрона, во всех окончаниях нейрона выделяется один и тот же медиатор. В настоящее время доказано, что этот принцип касается только пресинаптического единства нейрона. Эффекты, которые вызываются данным медиатором, могут быть различны и зависят от клеток-мишеней (в данном случае постсинаптических нейронов). Знак синаптичес-кого действия — повышение постсинаптического потенциала или его падение — определяется не медиатором, а свойствами рецепторов на постсинаптической клетке.
Постсинаптические рецепторы одного пресинаптического нейрона могут фармакологически различаться и контролировать разные ионные каналы. Одна постсинаптическая клетка может иметь более одного типа рецепторов для данного медиатора, и каждый из этих рецепторов контролирует отличный от других механизм ионной проводимости.
Кроме нейронов, суммирующих и передающих информацию к другим клеткам, описаны так называемые пейсмекерные нейроны, способные самостоятельно генерировать электрические импульсы (Alving, 1968). Активность таких нейронов характеризуется синусоидальными колебаниями частотой 0,1-10 Гц и амплитудой 5-10 мВ. Эти нейроны при отсутствии любого внешнего воздействия обеспечивают периодическую генерацию ПД и передачу возбуждения другим нейронам.
Медиаторы
В начале XX века группа английских физиологов, возглавляемая Дж. Лэнг-ли, показала, что электрическая стимуляция вегетативных нервов вызывает изменения в органах, иннервируемых этими нервами. Оказалось также, что такие изменения можно вызвать инъекцией в организм экстрактов надпочечников. Дж. Лэнгли предположил, что клетки, иннервируемые вегетативными нервами, имеют две рецептивные субстанции — тормозную и возбуждающую.
На основании этих данных Т. Эллиот в 1905 г. выдвинул предположение, что импульсы в вегетативных нервах вызывают выделение адреналина. В 1921 г. австрийский ученый О. Леви обнаружил, что тормозное влияние блуждающего нерва на деятельность сердца опосредуется специфическим веществом, позднее идентифицированным как ацетилхолин. Г. Дейл привел веские аргументы в пользу того, что ацетилхолин является медиатором в вегетативных ганглиях и нервно-мышечных соединениях. Однако доказать наличие синаптической передачи с помощью медиатора, а не электрического потенциала стало возможным только в 50-х годах, когда исследователи начали использовать микроэлектроды и электронный микроскоп.
Все медиаторные соединения — это низкомолекулярные водорастворимые (дипольные) амины или аминокислоты и родственные им вещества. Ацетилхолин и катехоламины синтезируются из циркулирующих в крови предшественников, тогда как аминокислоты и пептиды в конечном счете образуются из глюкозы. Свидетельством консерватизма живой природы является то, что, несмотря на различие циркуляторных систем и метаболических путей, беспозвоночные и позвоночные животные в равной степени используют большинство общих медиаторов (табл. 1.3).
Число пептидов, для которых доказаны медиаторные свойства, постоянно растет. Многие из этих веществ содержат от 2 до 10 аминокислот, что соответствует размеру, с одной стороны, мелких аминокислотных медиаторов, с другой — гормонов. Обилие пептидов создает впечатление неоднородности этой группы веществ. В то же время нарастающая информация о их роли
| Таблица 1.3. Характеристика некоторых медиаторов Полное название Принятое Замечания сокращение Ацетилхолин АХ Медиатор парасимпатической периной системы, имеется также и в головном мозге БИОГЕННЫЕ АМИНЫ Группа медиаторов, содержащих аминную Моноамины группу Серотин (5-окситриптамин) Кроме головного мозга присутствует в больших концентрациях в кишечнике и тромбоцитах Гистамин Кроме головного мозга в больших концентрациях находится в лимфоцитах Катехоламины КА В эту группу объединены нейромедиаторы — производные пирокатехина (дофамин, норадреналин, адреналин) Дофамин Возможное название допамин Норадреналин НА В англоязычной литературе называется норэпинефрин Адреналин А В англоязычной литературе называется эпинефрин АМИНОКИСЛОТЫ Глутамат Глу Возможно, самый распространенный возбуждающий медиатор Глицин Гли Основной ингибиторный медиатор ствола мозга и спинного мозга Аспартат Асп Возбуждающий медиатор Гамма-аминомасляная ГАМК Ингибиторный медиатор головного мозга кислота Алании Ала Ингибиторный медиатор ПЕПТИДЫ Цепи аминокислот Опиоидные пептиды Природные лиганды морфиновых рецепторов (эндогенные опиаты): энкефалины, эндорфины, динорфины Вещество Р Медиатор нервов, несущих информацию о боли Вазопрессин Гипофизарный гормон Окситоцин Гипофизарный гормон Энкефалины Гипофизарные гормоны Эндорфины Динорфины Недавно открытый класс опиатов ДРУГИЕ МЕДИАТОРЫ Аденозин Нуклеотид Реализуется в некоторых связях нерв — гладкая мышца |
в организме позволяет увидеть универсальные принципы их действия. Предполагается, что нейроэндокринные клетки, секретирующие пептиды, первыми появились в эволюции примитивных нервных систем. По-видимому, нейропептиды, производимые ими, достаточно консервативны, поскольку, как уже упоминалось, одинаковые вещества или близкие последовательности аминокислот обнаружены у филогенетически различных ветвей животных — беспозвоночных и позвоночных. Многие из них найдены не только в мозге, но и, например, в кишечнике. Есть предположение, что все пептид-эргические клетки связаны общностью эмбрионального происхождения. Пептиды по сравнению с другими медиаторами оказывают свое действие в чрезвычайно низких концентрациях.
С тех пор как в 1921 г. был идентифицирован первый медиатор, число их в арсенале науки постоянно увеличивается и в настоящее время составляет около 50. Многие биологически активные вещества имеют сходную с ними структуру. Они могут усиливать действие медиаторов (такие вещества называются агонистами) или подавлять их активность (антагонисты). Например, лекарственные препараты, снимающие тревогу (седуксен и др.), усиливают действие тормозного нейромедиатора — гамма-аминомасляной кислоты. Антидепрессанты (например, прозак) являются агонистами серо-тонина. Кокаин усиливает действие дофамина. Он связывается с белком, удаляющим дофамин из места его активности, тем самым увеличивая время его действия. Никотин активирует рецепторы ацетилхолина. Энкефалины и эндорфины являются природными лигандами морфиновых рецепторов: в норме именно они связываются с рецепторами, с которыми в особых условиях — при употреблении наркотика — взаимодействует морфин.
Важным звеном в нервном пути, который служит мишенью всех наркотиков — амфетамина, никотина, алкоголя и опиатов, — является небольшая часть базальных ганглиев, называемая nucleus accumbens (прилежащее ядро). Антипсихотические препараты (нейролептики) предотвращают связывание дофамина с его рецепторами. Содержащие дофамин нейроны, находящиеся в области вентральной покрышки среднего мозга, посылают свои аксоны в префронтальную кору и базальные ганглии, которые участвуют в двигательном контроле. Это обусловливает побочное действие длительно применяющихся нейролептиков, связанное с развитием дискинезий.
В настоящее время описан еще один класс посредников, имеющих чрезвычайно малые размеры молекул. К ним относят оксид азота (NO) и оксид углерода, или угарный газ (СО). Оксид азота опосредует действие ацетилхолина при расширении сосудов, в том числе сердечной мышцы. Именно он является активным компонентом нитроглицерина, используемого для расслабления резко суженных венечных сосудов сердца при стенокардии (грудной жабе). Этот посредник обнаружен в 2% клеток мозга. Он крайне токсичен, поэтому используется макрофагами (одним из видов клеток иммунной системы) для уничтожения бактерий, проникающих в организм. Второй посредник — угарный газ — не менее токсичен и опасен для человека. В мозге он выполняет важную роль, активируя вторичный (клеточный) посредник — цГМФ.
Белки, служащие рецепторами нейромедиаторов, можно разделить на два класса в зависимости от механизма их действия. К одному классу относятся белки ионных каналов, меняющие свою форму и открывающие каналы, по которым проходят ионы. Рецепторы другого класса располагаются по соседству с мембранными G-белками, разрывающими богатую энергией фосфатную связь в молекуле гуанозинтрифосфата, что инициирует каскад биохимических процессов, ведущих к специфической клеточной реакции через вторичные (клеточные) посредники. Эффекты, производимые этими белками, характеризуются медленным началом действия и большей продолжительностью, по сравнению с реакциями, связанными с открытием ионных каналов.
Функции нейрона
В настоящее время можно говорить о наличии трех основных функций нейрона. Наиболее распространенной является суммация возбуждающих и тормозных синаптических потенциалов и передача возбуждения следующему нейрону.
Описаны нейроны (прежде всего нейроны гипоталамуса), обладающие секреторной функцией. Они синтезируют биологически активные вещества — статины и либерины — и выделяют их в кровеносные сосуды воротной системы гипоталамуса. С током крови эти вещества попадают в переднюю долю гипофиза и способствуют реализации или накоплению его гормонов.
Наконец, существуют нейроны, обладающие спонтанной ауторитмичес-кой активностью. Их называют пейсмекерами, или водителями ритма. Эндогенные процессы подобных нейронов приводят к периодическому изменению ионной проницаемости мембраны и генерированию ПД. Взаимодействуя с другими клетками, они синхронизируют активность этих клеток.
Таблица ].4. Свойства различных нервных волокон теплокровных (Бабский, 1972).
| Длитель- | Длитель- | |||||
| Тип волокна | Диаметр волокна, (мк) | Скорость проведения, (м/сек) | Длительность ПД, (м/сек) | ность отрицательного следово-гоПД, | ность положительного следово-гоПД, | Функция |
| (м/сек) | (м/сек) | |||||
| Моторные волокна | ||||||
| А(а) | 12-22 | 70-120 | 0,4-0,5 | 12-20 | 40-60 | скелетных мышц, афферентные волокна от |
| мышечных рецепторов | ||||||
| А(Ь) | 8-12 | 40-70 | 0,4-0,6 | — | Афферентные волокна от рецепторов прикосновения | |
| Афферентные волокна от | ||||||
| рецепторов прикосновения и | ||||||
| A(g) | 4-8 | 15-40 | 0,5-0,7 | — | давления, афферентные | |
| волокна к мышечным | ||||||
| веретенам | ||||||
| Афферентные волокна от | ||||||
| A(D) | 1-4 | 5-15 | 0,6-1,0 | — | — | некоторых рецепторов тепла, |
| давления, боли | ||||||
| В | 1-3 | 3-14 | 1-2 | Отсутствует | 100-300 | Преганглионарные вегетативные волокна |
| Постганглионарные | ||||||
| вегетативные волокна, | ||||||
| С | 0,5-1,0 | 0,5-2 | 2,0 | 50-80 | 300-1000 | афферентные волокна от |
| некоторых рецепторов тепла, | ||||||
| давления, боли |
Типы нервных волокон
По скорости проведения импульса и строению нервные волокна разделяют на три группы — А, В и С. Волокна типа А делятся на 4 подгруппы: альфа-, бета-, гамма-, дельта-. Альфа-волокна имеют самый большой диаметр (12-22 мк) и обладают наибольшей скоростью проведения возбуждения (70-120 м/сек). Такие волокна проводят информацию к скелетным мышцам и от них в мозг, что позволяет человеку достаточно быстро приспосабливать положение своего тела к ситуации. Остальные нервные волокна имеют меньший диаметр (2-12 мк) и соответственно меньшую скорость проведения импульса. Они несут информацию отсенсорных органов. Гамма-волокна передают возбуждение от моторных нейронов спинного мозга к интрафузальным мышечным волокнам.
К волокнам типа В относятся миелинизированные волокна, преимуще-твенно преганглионарные, распространенные в автономной нервной системе. Скорость проведения информации по ним составляет 3-14 м/сек.
Волокна типа С — это немиелинизированные волокна, скорость проведения информации в них составляет 2-6 м/сек. Большинство из них являются
постганглионарными волокнами автономной нервной системы. Эти волокна проводят также сигналы о боли, тепле, давлении (табл. 1.4) (Бабский, 1972).
Словарь
Вегетативная нервная система —
иннервирует гладкую мускулатуру и управляет деятельностью внутренних органов, поэтому ее также называют висцеральной.
Везикулы —
особые пузырьки, расположенные в аксонной терминали, в которых хранится медиатор.
Гематоэщефалический барьер —
уникальный защитный слой между нейроном и кровеносным сосудом, образованный гли-альными клетками.
/ЛИЯ —
функционально разнообразные клетки мозга, способные делиться, но не участвующие в процессе передачи информации.
Модуль —
объединение нейронов коры, несущих одну функцию, в виде колонок. Модуль может вклю-
чать более 100 тыс. клеток, огромное большинство которых образует локальные нейронные сети, выполняющие ту или иную функцию.
Нейрон —
основная функциональная клетка мозга, участвующая в передаче и хранении информации. Это поляризованная клетка, которая с помощью сильно разветвленных многочисленных отростков — дендритов — получает сигналы и через длинный неразветвленный отросток аксон посылает информацию другой клетке.
|
из
5.00
|
Обсуждение в статье: Синаптическая передача информации |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы