 |
Главная |


Глава 9. ВНЕШНЕЕ ДЫХАНИЕ
|
из
5.00
|
Основные вопросы: Понятие о дыхании и его основные этапы. Анатомическое строение дыхательного тракта человека. Основные фазы дыхательного цикла. Механизм вдоха и выдоха. Легочные объемы. Состав вдыхаемого, выдыхаемого и альвеолярного воздуха. Минутный объем дыхания и минутная вентиляция легких. Анатомическое и физиологическое дыхательное мертвое пространство. Типы легочной вентиляции. Обмен дыхательных газов в организме.
Дыхание – это совокупность процессов, обеспечивающих поступление во внутреннюю среду О2, который используется для окисления органических веществ, и удаление из организма СО2, который образуется в результате тканевого метаболизма.
Выделяют три основных этапа дыхания:
1) внешнее дыхание;
2) транспорт дыхательных газов;
3) внутреннее дыхание.
I этап - внешнее дыхание - это проведение воздуха, обновление газовой смеси и обмен газов в дыхательном тракте.
Дыхательный тракт человека состоит из воздухоносных путей и респираторной зоны органа дыхания – легких (рис.45).

Рис.45. Дыхательный тракт человека
Респираторная зона легких представлена тонкостенными легочными мешочками – альвеолами, которые окружены густой сетью кровеносных капилляров.
Воздухопроводящие пути подразделяют на верхние и нижние. К верхним относятся: носовые ходы, полость рта, носоглотка, а к нижним - гортань, трахея, бронхи и бронхиолы.
Основные функции воздухоносных путей:
1) проведение воздуха в респираторную зону;
2) согревание воздуха;
3) увлажнение воздуха;
4) очищение воздуха.
Трахею и следующие за ней 16 первых разветвлений (генераций) бронхов и бронхиол, относят к кондуктивной (проводящей) зоне легких. В этой зоне отсутствует контакт между воздухом и кровеносными легочными капиллярами, а значит, не происходит газообмен.
Последующие три генерации бронхиол (17-19) составляют переходную (транзиторную) зону, где наряду с проведением воздуха начинает осуществляться газообмен. Последние четыре генерации бронхиол (с 20 по 23) представляют собой альвеолярные ходы, переходящие в альвеолы (рис.46).
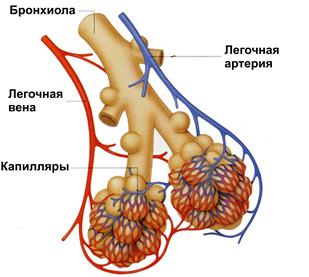
Рис.46. Легочные альвеолы
Возвратно-поступательные перемещения воздушных масс в респираторном тракте, обеспечивающее обновление газового состава воздуха в альвеолах, называют легочной вентиляцией. Обмен дыхательных газов (О2, СО2) между альвеолами и кровью легочных капилляров – легочная диффузия.
II этап - транспорт газов - перенос кровью кислорода от легких к тканям, а углекислого газа от тканей к легким.
III этап - внутреннее (тканевое) дыхание, включает в себя:
1) обмен дыхательных газов между кровью тканевых капилляров и клетками;
2) биохимические процессы окисления кислородом органических веществ в клетках.
Благодаря тканевому дыханию клетки получают энергию, необходимую для жизнедеятельности, и освобождаются от углекислоты, которая накапливается в процессе метаболизма.
Вентиляция легких происходит вследствие периодических изменений объема грудной полости в процессе дыхательного цикла.
Полный дыхательный цикл состоит из трех основных фаз:
1) фаза вдоха (инспирация);
2) фаза выдоха (экспирация);
3) дыхательная пауза.
Изменения объема грудной полости в процессе дыхательного цикла обусловлены ритмическими сокращениями и расслаблениями дыхательных мышц. Они подразделяются на инспираторные и экспираторные.
Различают основные и вспомогательные инспираторные мышцы.
Основные инспираторные мышцы:
1) диафрагма;
2) наружные межреберные мышцы.
Главная инспираторная мышца – куполообразная диафрагма, отделяющая грудную полость от брюшной. При сокращении мышечных волокон купол диафрагмы уплощается, что ведет к увеличению вертикального размера грудной полости. Сокращение наружных межреберных мышц ведет к приподниманию ребер и увеличению передне-заднего и бокового объема грудной клетки (рис.47).
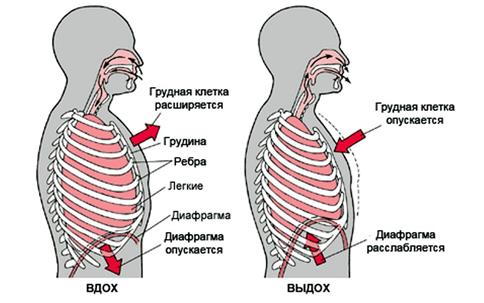
Рис.47. Механизм вдоха и выдоха
Во время вдоха инспираторные мышцы преодолевают эластическое сопротивление тканей грудной клетки, брюшной стенки, органов брюшной полости и легких.
Эластическое сопротивление легких – эластическая тяга, обусловлено:
1) наличием большого числа эластических волокон в тканях легких;
2) упруго-растяжимыми свойствами легочных альвеол;
3) сурфактантом.
Около 2/3 от общей величины эластической тяги легких создается сурфактантом - комплексом веществ липопротеидной природы, которые выстилают изнутри легочные альвеолы.
Сурфактант обеспечивает:
1) стабилизацию сферической формы легочных альвеол;
2) противодействие перерастяжению альвеол при вдохе;
3) противодействие спадению альвеол при выдохе;
4) очищение внутренней поверхности альвеол.
Легкие находятся внутри грудной клетки и отделены от ее стенок плевральной щелью – узкой, герметически замкнутой полостью. Давление в плевральной полости ниже атмосферного.
Спокойный вдох начинается с сокращения основных инспираторных мышц. При сокращении инспираторных мышц грудная клетка расширяется во всех направлениях. Объем грудной полости увеличивается.
Увеличение объема грудной полости последовательно вызывает:
1) снижение давления в плевральной щели до -6 -9 мм рт. ст. (по сравнению с атмосферным);
2) расширение легких;
3) снижение внутрилегочного давления до -2 мм рт. ст.;
4) поступление воздуха в легкие по градиенту между атмосферным и альвеолярным давлением.
Cпокойный выдох начинается с расслабления основных инспираторных мышц. Это ведет к уменьшению объема грудной полости.
Уменьшение объема грудной полости последовательно вызывает:
1) повышение давления в плевральной щели с -6 -9 мм рт. ст. до -3 мм рт. ст.;
2) уменьшение объема легких за счет их эластической тяги;
3) повышение внутрилегочного давления до +2 мм рт. ст. (по сравнению с атмосферным);
4) выход воздуха из легких в атмосферу по градиенту давления.
Таким образом, непосредственной причиной поступления воздуха в легкие во время вдоха и его выход в атмосферу во время выдоха является градиент между внутрилегочным и атмосферным давлением. Спокойный выдох происходит пассивно вследствие расслабления инспираторных мышц.
Экспираторные и вспомогательные инспираторные мышцы обеспечивают форсированную вентиляцию легких.
Объем воздуха, который находится в легких после максимально глубокого вдоха, называется общей емкостью легких (ОЕЛ). У взрослого человека ОЕЛ составляет от 4200 до 6000 мл и состоит из двух частей:
1) жизненной емкости легких (ЖЕЛ) - 3500-5000 мл;
2) остаточного объема легких (ООЛ) - 1000-1200 мл.
Остаточный объем - это количество воздуха, которое остается в легких даже после максимально глубокого выдоха.
Жизненная емкость легких - это объем воздуха, который можно максимально выдохнуть после максимально глубокого вдоха.
Жизненная емкость легких состоит из трех частей:
1) дыхательный объем (ДО) - 400-500 мл;
2) резервный объем вдоха - около 2500 мл;
3) резервный объем выдоха - около 1500 мл.
Дыхательный объем - это количество воздуха, который вдыхается или выдыхается во время каждого дыхательного цикла при спокойном дыхании.
Резервный объем вдоха - это максимальное количество воздуха, которое можно дополнительно вдохнуть после спокойного вдоха.
Резервный объем выдоха - это максимальное количество воздуха, которое можно дополнительно выдохнуть после спокойного выдоха.
Резервный объем выдоха и остаточный объем составляют функциональную остаточную емкость (ФОЕ) - объем воздуха, остающееся в легких после спокойного выдоха (2000-2500 мл).
Легочная вентиляция характеризуется минутным объемом дыхания (МОД) - количеством воздуха, которое вдыхается или выдыхается за 1 мин. МОД зависит от величины дыхательного объема и частоты дыхания (ЧД): МОД = ДО х ЧД. У взрослого человека частота дыхания в покое составляет 12-16 дыхательных циклов в мин. Поэтому МОД колеблется от 6 до 10 л/мин. При физической нагрузке или эмоциональном возбуждении МОД может повышаться до 100-120 л/мин за счет увеличения ДО и ЧД.
В обычных условиях человек дышит атмосферным воздухом, в составе которого содержится: О2 - 21%, СО2 - 0,03%, N2 - 79%. В выдыхаемом воздухе: О2 - 16,0%, СО2 - 4%, N2 -79,7%. В альвеолярном воздухе: О2 - 14,0%, СО2 - 5,5%, N2 - 80%.
Различие в составе выдыхаемого и альвеолярного воздуха обусловлено смешиванием альвеолярного газа с воздухом дыхательного мертвого пространства. Различают анатомическое и физиологическое мертвое пространство.
Физиологическое дыхательное мертвое пространство (ФДМП) - это объем всех участков дыхательного тракта, в которых не происходит газообмен с кровью легочных капилляров.
К физиологическому мертвому пространству относятся объем воздухоносных путей, а также объемы невентилируемых или неперфузируемых кровью альвеол. Частью ФДМП является анатомическое дыхательное мертвое пространство - объем воздуха, заполняющего воздухопроводящие пути (от полости носа до бронхиол).
Объем анатомического мертвого пространства составляет примерно 150 мл. Поэтому, из 500 мл вдыхаемого воздуха только 350 мл поступает в альвеолы респираторной зоны легких. Здесь этот воздух смешивается с альвеолярным газом, который составляет функциональную остаточную емкость легких. Величина ФОЕ около 2500 мл, поэтому, при каждом спокойном вдохе обновляется только 1/7 часть альвеолярного воздуха (2500 мл:350 мл).
Воздух, заполняющий мертвое пространство, выполняет функцию буфера, который сглаживает колебания состава альвеолярной смеси газов в ходе дыхательного цикла. Это обеспечивает относительное постоянство состава воздуха, в легочных альвеолах, и непрерывность легочной диффузии независимо от фазы дыхательного цикла.
Количество воздуха, который участвует в обмене газов между альвеолами и кровью легочных капилляров за 1 мин, называется минутной вентиляцией легких (МВЛ). МВЛ = (ДО - ФДМП) х ЧД.
Различают десять основных типов вентиляции легких:
1) нормовентиляция;
2) гипервентиляция;
3) гиповентиляция,
4) эйпноэ;
5) гиперпноэ;
6) гипопноэ;
7) тахипноэ;
8) брадипноэ;
9) апноэ;
10) диспноэ.
Нормовентиляция - это уровень газообмена в легких, который соответствует метаболическим потребностям организма. Такой уровень газообмена в покое обеспечивается эйпноэ – нормальным спокойным дыханием.
Гипервентиляция – это такой уровень газообмена в легких, который превышает метаболические потребности организма. Причиной гипервентиляции может быть гиперпноэ - увеличение глубины дыхания выше нормы, и тахипноэ - увеличение частоты дыхания выше нормы.
Гиповентиляция - это газообмен в легких, который не достаточен для обеспечения метаболических потребностей организма. Причиной гипервентиляции может быть гипопноэ - уменьшение глубины дыхания ниже нормы, и брадипноэ - уменьшение частоты дыхания ниже нормы.
Диспноэ (одышка) - это затрудненность дыхания, которая сопровождаются неприятными субъективными ощущениями. Крайним проявлением диспноэ может быть апноэ - остановка дыхания.
Перемещение смеси газов в воздухоносных путях происходит в результате конвекции и диффузии. Конвекция - движение смеси газов по градиенту их общего давления. В ходе ветвления воздухоносных путей их суммарное сечение значительно возрастает. Это ведет к снижению линейной скорости потока вдыхаемого воздуха со 100 см/с до 0,02 см/с. В этих условиях к конвекции присоединяется диффузионный обмен.
Диффузия газа - это пассивное движение молекул газа из области большего парциального давления или напряжения в зону меньшего. Парциальное давление газа - это часть общего давления, которая приходится на какой-либо газ, смешанный с другими газами. Парциальное давление газа, растворенного в жидкости, называют напряжением газа.
Перенос О2 из альвеолярного газа в венозную кровь и СО2 из крови в легочные альвеолы происходит пассивно за счет разности парциального давления и напряжения этих газов по обе стороны аэрогематического барьера.
Аэрогематический барьер образован альвеолокапиллярной мембраной, которая включает в себя слой сурфактанта, а также альвеолярный эпителий, эндотелий кровеносного капилляра и их базальные мембраны (рис.48).

Рис.48. Аэрогематический барьер
Парциальное давление О2 в альвеолярном воздухе 100 мм рт. ст. Напряжение О2 в венозной крови легочных капилляров 40 мм рт. ст. Градиент давления, составляющий 60 мм рт. ст., направлен из альвеол в кровь.
Парциальное давление СО2 в альвеолярном воздухе 40 мм рт. ст. Напряжение СО2 в венозной крови легочных капилляров 46 мм рт. ст. Градиент давления, составляющий 6 мм рт. ст., направлен из крови в альвеолы.
Время протекания крови через легочные капилляры составляет около 0,75 с. Этого достаточно для практически полного выравнивания давления газов по обе стороны аэрогематического барьера. При этом кислород растворяется в венозной крови, а двуокись углерода переходит в легочные альвеолы. Поэтому венозная кровь превращается здесь в артериальную.
Напряжение О2 в артериальной крови 100 мм рт. ст., а в тканях организма менее 40 мм рт. ст. Градиент давления, составляющий более 60 мм рт. ст., направлен из артериальной крови в ткани.
Напряжение СО2 в артериальной крови 40 мм рт. ст., а в тканях - около 60 мм рт. ст. Градиент давления, составляющий 20 мм рт. ст., направлен из тканей в кровь. Благодаря этому артериальная кровь в тканевых капиллярах превращается в венозную.
Таким образом, звенья газотранспортной системы характеризуются встречными потоками дыхательных газов: О2 перемещается из атмосферы к тканям, а СО2 - в обратном направлении.
Глава 10. ТРАНСПОРТ ДЫХАТЕЛЬНЫХ ГАЗОВ КРОВЬЮ.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Основные вопросы: Дыхательный центр. Особенности автоматии дыхательного центра. Механизмы саморегуляции дыхания. Механо- и хеморецепторные механизмы регуляции дыхания. Гуморальная регуляция дыхания. Специфические и неспецифические регуляторы дыхания, дыхание при физической работе.
Пути транспорта О2 кровью. Кислородная емкость крови. Кривая диссоциации оксигемоглобина. Факторы, влияющие на образование и диссоциацию оксигемоглобина. Механизмы транспорта СО2 кровью, значение карбоангидразы эритроцитов.
Оптимальное содержание газов в крови достигается регуляцией объема легочной вентиляции в зависимости от текущих потребностей метаболизма.
Регуляция внешнего дыхания - это физиологический процесс управления вентиляцией легких, который направлен на достижение полезного приспособительного результата - оптимального газового состава внутренней среды организма.
Координированные сокращения дыхательных мышц, обеспечивающих легочную вентиляцию, управляются нейронами дыхательного центра.
Дыхательный центр - это совокупность нейронов, расположенных на разных этажах ЦНС и обеспечивающих координированную, ритмическую сократительную деятельность дыхательных мышц в процессе приспособления дыхания к изменяющимся условиям внешней и внутренней среды.
В регуляции дыхания участвуют:
1) спинной мозг;
2) задний мозг;
3) кора больших полушарий головного мозга.
Жизненно важная часть дыхательного центра расположена в заднем мозге (в продолговатом мозге и варолиевом мосту). Она включает в себя три отдела:
1) инспираторный;
2) экспираторный;
3) пневмотаксический.
Инспираторный отдел состоит из нейронов, генерирующих ПД в фазу вдоха. Экспираторный отдел включает в себя нервные клетки, которые возбуждаются во время выдоха. Инспираторный и экспираторный отделы дыхательного центра находятся в реципрокных отношениях: при возбуждении инспираторных нейронов угнетается экспираторный отдел и наоборот. Нейроны пневмотаксического отдела проявляют максимум разрядной электрической активности при смене фаз дыхательного цикла. Они обеспечивают плавный переход от вдоха к выдоху.
Нейроны жизненно важной части дыхательного центра образуют микрокомплексы, обладающие автоматией, т.е. способностью самопроизвольно генерировать ПД.
Имеется четыре основные особенности автоматии дыхательных нейронов:
1. Автоматией обладают микрокомплексы, состоящие из инспираторных и экспираторных дыхательных нейронов продолговатого мозга. Эти нейроны образуют ритмозадающие группы, которые являются генератором дыхательной периодики.
2. Автоматия микрокомплексов дыхательных нейронов поддерживается афферентными сигналами от периферических (локализуются в сосудистом кровеносном русле) и центральных хеморецепторов (располагаются в ЦНС). Сигналы, поступающие от центральных и периферических рецепторов, являются необходимым условием периодической активности дыхательного центра и механизмом приспособления вентиляции легких газовому составу крови.
3. Дыхательные нейроны продолговатого мозга контролируются пневмотаксическим отделом дыхательного центра и подкорковыми ядрами.
Пневмотаксический центр не является генератором дыхательной периодики, но постоянно участвует в регуляции частоты дыхания за счет увеличения скорости развития инспираторной активности, активации механизма переключения с инспирации на экспирацию.
4. Автоматическая разрядная деятельность дыхательных микрокомплексов подчиняется сознательному контролю. Особую роль такая регуляция дыхания играет у человека, что связано с участием дыхательного аппарата в речевой функции.
Человек может произвольно менять не только частоту и глубину дыхания, но и временно задерживать его. Однако длительность произвольного апноэ ограничена: как только напряжение СО2 в крови достигает критического уровня, сильное возбуждение хеморецепторов выводит ритмозадающие микрокомплексы из-под произвольного контроля и, независимо от желания человека, дыхательные движения возобновляются.
Дыхательные микрокомплексы управляют сократительной активностью дыхательных мышц через a -инспираторные нейроны, которые локализуются в продолговатом мозге. Периодическое автоматическое возбуждение a-инспираторных нейронов ведет к периодической активации дыхательных a -мотонейронов, которые расположены в шейном и грудном отделе спинного мозга. Аксоны дыхательных a-мотонейронов, которые расположены во 2-5 шейных сегментах спинного мозга формируют диафрагмальный нерв, иннервирующий основную дыхательную мышцу. Аксоны дыхательных a-мотонейронов, локализующихся в грудных сегментах спинного мозга, образуют межреберные нервы иннервирующие косые межреберные мышцы (рис.49).

Рис.49. Схема иннервации диафрагмы
1) a-инспираторный нейрон (Ia), 2) дыхательные a-мотонейроны 2-5 шейных сегментов спинного мозга, 3) диафрагмальный нерв, 4) мышечные волокна диафрагмы.
Генерация дыхательными a-мотонейронами спинного мозга группы (пачки) ПД вызывает сокращение инспираторных мышц. Прекращение электрической разрядной деятельности дыхательных a-мотонейронов приводит к расслаблению инспираторных мышц.
Глубина и частота дыхание зависят от параметров разрядной деятельности дыхательных a-мотонейронов. Частота дыхания зависит от частоты генерации пачек ПД (от межпачечного интервала). Чем больше частота следования пачек, тем больше частота дыхания.
Глубина дыхания определяется двумя факторами:
1) частотой ПД в пачках;
2) количеством вовлеченных в процесс возбуждения моторных единиц.
Чем больше частота ПД в пачке (чем больше разрядов за единицу времени), тем больше амплитуда сокращения дыхательной мышцы, а значит и глубина вдоха.
Каждый дыхательный a-мотонейрон иннервирует не одно мышечное волокно, а целую группу. Тело нейрона, его аксон, коллатерали (ответвления) и группа мышечных волокон, которые он иннервирует, составляют моторную единицу (рис.50).

Рис.50. Схема моторной единицы
1) тело мотонейрона, 2) аксон, 3) коллатерали, 4) мышечные волокна.
Чем больше возбуждено моторных единиц, тем больше мышечных волокон вовлечено в сократительную реакцию, больше амплитуда мышечного сокращения, а значит и глубина дыхания.
Механизмы саморегуляцииглубины и частоты дыхания подразделяются на нервные и гуморальные.
Нервные механизмы связаны с рефлекторными реакциями внешнего дыхания, которые возникают при раздражении специализированных периферических хеморецепторов и механорецепторов.
В зависимости от вида рецепторов различают механорецепторный и хеморецепторный механизмы рефлекторной регуляции легочной вентиляции.
Механорецепторная регуляция связана с раздражением механорецепторов, которые реагируют на растяжение воздухоносных путей легких. В экспериментах на животных Герингом и Брейером обнаружено преждевременное прекращение вдоха при искуственном раздувании легких во время инспирации (рефлекс Геринга-Брейера).
Рефлекс Геринга-Брейера возникает вследствие раздражения рецепторов растяжения, от которых по афферентным волокнам блуждающего нерва управляющие сигналы поступают в центр регуляции дыхания. В естественных условиях возбуждение рецепторов растяжения легких, нарастая в ходе вдоха, рефлекторно приводит к плавной смене инспирации экспирацией.
При выключении афферентного звена путем перерезки блуждающих нервов (ваготомии) вдохи становятся углубленными и затянутыми. Если же ваготомию сочетать с разрушением пневмотаксического центра, то наблюдается айпнезис: дыхательные движения останавливаются на вдохе, который изредка прерывается короткими экспирациями.
Хеморецепторная регуляции обусловлена раздражением периферических хеморецепторов, которые локализуются в двух рефлексогенных зонах: Геринга и Циона-Людвига. При этом ведущее значение имеет рефлексогенная зона Геринга.
Каротидное тельце зоны Геринга расположено в области бифуркации сонной артерии. Оно содержит сложно устроенный рецепторный аппарат, который реагирует на изменения дыхательных показателей артериальной крови: гиперкапнию, гипоксемию и ацидоз. Все три фактора вызывают возбуждение каротидных хеморецепторов, что ведет к активации дыхательного центра и рефлекторному увеличению частоты и глубины дыхания.
Главным стимулом, управляющим функцией легочной вентиляции, является гиперкапния. Однако наиболее значима чувствительность периферических хеморецепторов зоны Геринга к гипоксемии. Они являются единственными в организме сигнализаторами о недостатке О2.
Выраженное влияние на деятельность дыхательного центра оказывает гуморальная стимуляция центральных хеморецепторов. Наибольшее их скопление находится на переднебоковых поверхностях продолговатого мозга. В обычных условиях центральные хеморецепторы постоянно активируются ионами водорода спинномозговой и межклеточной жидкости ствола мозга. Концентрация ионов водорода зависит от напряжения СО2 в крови.
Спинномозговая жидкость отделена от крови гематоэнцефалическим барьером, который относительно непроницаем для ионов Н+ и НСО3-, но свободно пропускает молекулярный СО2. При повышении напряжения СО2 в крови он проникает в спинномозговую жидкость. В результате реакции гидратации с помощью карбоангидразы из СО2 образуется Н2СО3, которая диссоциирует на катионы Н+ и анионы НСО3-. Ионы Н+ непосредственно стимулируют центральные хеморецепторы продолговатого мозга. Вследствие возбуждения центральных хеморецепторов повышается разрядная активность нейронов дыхательного ритмообразующего комплекса. В результате этого учащается и углубляется дыхание, увеличивается легочная вентиляция.
Напротив, снижение напряжения СО2 и подщелачивание межклеточной жидкости ведет к угнетению активности дыхательного центра вплоть до остановки дыхания.
Сильным специфическим стимулятором дыхания является физическая работа. Частота и глубина дыхания увеличивается непосредственно после начала движений. После окончания работы уровень легочной вентиляции в течение более или менее продолжительного времени, в зависимости от тяжести предшествовавшей работы, остается повышенным. В этот период хеморецепторы стимулируются циркулирующими в крови недоокисленными продуктами обмена.
Основными неспецифическими регуляторами дыхания являются:
1) температурные воздействия;
2) эмоционально значимые и болевые стимулы;
3) повышение артериального давления.
Гипертермия или умеренная гипотермия сопровождаются увеличением вентиляции легких. Однако резкое охлаждение приводит к угнетению дыхательных центров. За счет торможения инспираторных и экспираторных нейронов угнетается дыхание и при повышении артериального давления.
Болевые и эмоционально значимые стимулы активируют дыхание.
|
из
5.00
|
Обсуждение в статье: Глава 9. ВНЕШНЕЕ ДЫХАНИЕ |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы