 |
Главная |


Рост и развитие растений. Общие закономерности роста и развития клеток, органов и целостного растительного организма. Влияние внешних факторов на процессы роста и развития.
|
из
5.00
|
В основе роста и развития целого организма и отдельной клетки лежит обмен веществ. В процессе жизни каждого организма происходят постоянные качественные и количественные изменения, прерываемые лишь периодами относительного покоя.
Необратимое количественное увеличение структур, объема и массы живого тела и его частей получило название роста. Развитие - это качественные изменения организма и его составляющих. Рост и развитие тесно связаны между собой, как правило, протекают параллельно, но не сводимы друг к другу. Оба процесса регулируются на клеточном уровне.
Рост отдельных органов и всего организма слагается из роста его клеток. Основные этапы роста, а также и развития на клеточном уровне - деление клеток и их растяжение, т.е. увеличение размеров в длину. Постепенное увеличение линейных размеров, объема и массы клеток - важнейшие показатели роста. В многоклеточных организмах одним из показателей роста является увеличение числа клеток в результате клеточного деления.
Растительная клетка способна к росту растяжением, чему содействуют особенности строения ее стенки. Длительность роста растяжением клеток различных тканей неодинакова. У части тканей, стенки которых способны к вторичным изменениям, рост растяжением на определенном этапе прекращается и наступает вторая фаза роста, при которой рост осуществляется путем наложения новых слоев на первичную оболочку или внедрением в нее.
Особенности роста различны у разных систематических групп организмов. У высших растений рост тесно связан с деятельностью меристем. Рост, так же как и развитие, контролируется фитогормонами. Помимо влияния фитогормонов на рост и развитие растения, заметное воздействие оказывают факторы среды, особенно свет, тепло и влага. Комплекс этих факторов и фитогормонов действует либо независимо, либо взаимодействуя друг с другом. Интенсивность роста существенным образом связана с питанием растений, особенно с азотным и фосфорным.
Типы роста различных органов определяются характером расположения меристем. Стебли и корни растут верхушками, т.е. имеют апикальный рост. Зона нарастания листьев часто находится у их основания, и они имеют базальный рост. Нередко характер роста органа зависит от видовой специфичности. У злаков, например, рост стебля осуществляется у основания междоузлий , когда преобладает интеркалярный рост. Важная особенность роста растений - его ритмичность, т.е. чередование процессов интенсивного и замедленного роста. Она зависит не только от изменений внешних факторов среды, но и контролируется внутренними факторами (эндогенно), закрепленными генетически в процессе эволюции.
В целом рост растения складывается из четырех фаз: начальной, интенсивного роста, замедления роста и стабильного состояния. Это связано с особенностями различных стадий онтогенеза , т.е. индивидуального развития растений.
Так, переход растения к репродуктивному состоянию обычно сопровождается ослаблением активности меристем . Процессы роста могут прерываться продолжительными периодами торможения, наступление которых в северных широтах связано с концом лета и приближением зимы. Иногда у растений наблюдается как бы остановка роста - состояние покоя. Покой у растений - это такое физиологическое состояние, при котором резко снижаются скорость роста и интенсивность обмена веществ . Оно возникло в ходе эволюции как приспособление для переживания неблагоприятных условий среды в различные периоды жизненного цикла или сезона года. Покоящееся растение значительно более устойчиво к морозам, жаре, засухе. В состоянии покоя могут находиться целые растения (зимой или во время засухи), их семена, почки, клубни, корневища, луковицы, споры и др. Семена многих растений способны к длительному покою, обусловливающему их надежную сохранность в почве. Известен случай развития нормального растения из семени одного из бобовых , пролежавшего в условиях вечной мерзлоты 10000 лет. В состоянии покоя находятся, например, клубни картофеля, благодаря чему они не прорастают некоторое время после уборки.
В понятие "развитие" вкладываются два смысла: индивидуальное развитие отдельного организма и развитие организмов в ходе эволюции. Индивидуальное развитие отдельного организма от рождения до смерти называется онтогенезом , а развитие организмов в ходе эволюции - филогенезом . Физиология растений занимается изучением развития главным образом в ходе онтогенеза.
Меристематические клетки полипотентны, т.е. дают начало клеткам, способным развиваться самыми различными путями.Переход меристематической клетки к росту растяжением сопровождается появлением в ней множества вакуолей и их слиянием в одну, вытягиванием стенок клеток в длину и поглощением вакуолями большого количества воды. Наиболее важный момент в развитии клеток высшего растения - их дифференцировка, т.е. возникновение структурной и функциональной разнокачественности. В результате дифференциации образуются специализированные клетки, присущие отдельным тканям. Дифференцировка осуществляется как во время растяжения, так и после окончания видимого роста клеток и определяется дифференциальной активностью генов. Ход дифференцировки, как и рост, контролируется фитогормонами
(Ответ на вопрос искал в нете, так что за достоверностью и полностью не ко мне)
18. Почка – зачаточный побег. Строение почки, конус нарастания. Типы почек по положению: верхушечные и боковые. Почки вегетативные, цветочные и смешанные.
Почка – зачаточный, еще не развившийся побег. Внутри почки заключена меристематическая верхушка побега – апекс (конус нарастания), основная часть которого представляет собой точку роста. Почки обычно окружены специализированными почечными чешуями, представляющими видоизмененные листья. В этих случаях говорят о закрытых почках. Количество почечных чешуй у разных видов различно. Открытые почки специальных чешуй не имеют. Деятельность конуса нарастания обеспечивает формирование всех тканей и частей побега. Сам конус нарастания не обязательно имеет геометрическую форму конуса, нередко это плоская или даже вогнутая вершинная часть апекса. На апексе побега регулярно формируются выступы, так называемые листовые примордии. Они появляются акропетальной последовательности, т.е. от основания к вершине. Гладким у апекса остается лишь верхушка, которую собственно и называют точкой роста. Заложение метамеров происходит ритмично. Промежуток времени между вычленением на апексе двух последовательных метамеров называется пластохроном ( 2-4 суток у древесных). Апекс состоит из нескольких частей. Обычно самый наружный слой клеток апикальной меристемы становится протодермой, т.е. меристемой, дающей начало эпидерме будущего стебля и листа. Клетки внутренней части апекса способны делиться во всех направлениях и обеспечивает его объемный рост. У двудольных и голосеменных на границе внутренней и наружной частей апекса на уровне листовых зачатков закладывается так называемая периферическая меристема, или инициальное кольцо. Именно здесь закладываются тяжи относительно узких и длинных клеток прокамбия, дающего начало I проводящим тканям. Прокамбий распространяется по мере дальнейшего роста апекса и вглубь оформляющегося стебля – оси побега, образуя основу будущей проводящей системы побега. Помимо I меристем, существуют основные меристемы. Из них формируются паренхимные клетки I коры стебля и сердцевина.
Можно выделить 2 типа почек – вегетативные и генеративные. Из первых развиваются сугубо вегетативные побеги, из вторых – цветки и соцветия. Цветочная почка, из которой развивается отдельный цветок, называется бутон. Весьма обычны вегетативно – генеративные почки, в которых помимо ряда вегетативных метамеров, заложены соцветие или отдельный цветок. Вегетативные почки подразделяются на ряд типов по физиологической роли и положению на побеге или растении, однако их морфологическое устройство весьма сходно. За счет верхушечной почки побега осуществляется его рост и развитие. Боковые, или пазушные, почки дают побеги следующего порядка. Они могут быть одиночными или множественными и располагаются в этом случае весьма различно, но всегда в пазухе кроющего листа. Н месте кроющего листа после его опадения на стебле образуется различной формы листовой рубец. У оснований стволов деревьев легко заметить спящие почки. В естественных условиях из спящих почек образуются корневая и пневая поросль - после отмирания или рубки леса. Спящие почки формируют систему скелетных осей многих кустарников. Придаточные, или адвентивные почки формируются на всех частях и органах растения, включая корни. Пазушные почки формируются экзогенно, придаточные и спящие почки возникают эндогенно из камбия, перицикла. У многих растений из придаточных почек на корнях образуются корневые отпрыски. Почки, впадающие на зиму в покой – зимующие, а в климате, не имеющем выраженной зимы, - покоящимися. Такие почки при возобновлении активного роста дают новые годичные или элементарные побеги. Почки, лишенные периода покоя, - почки обогащения, которые дают побеги обогащения. Побеги обогащения характерны для фасоли, настурции, погремка, мокрицы. Такие побеги существенно увеличивают фотосинтезирующую поверхность растения или становятся побегами, несущими боковые соцветия. За счет деятельности боковых и придаточных почек происходит постоянное нарастание системы побегов многолетних растений.
19. Побег – вегетативный орган растений. Метамерия побегов. Формирование побегов в онтогенезе.Укороченные и удлиненные побеги. Типы ветвления побегов. Листорасположение и его закономерности.
Побег – вегетативный орган растения, который обычно располагается в воздушной среде и выполняет функцию фотосинтеза. Исключение составляют видоизмененные подземные побеги (корневища, луковицы, клубни), которые располагаются в почве, в них накапливается запас продуктов фотосинтеза, эти побеги обеспечивают вегетативное размножение и захват территории. В отличие от корня, который имеет зональное строение, побег имеет метамерное строение, т.е. побег представляет собой закономерное чередование метамеров, т.е. узлов с располагающимися выше междоузлиями. Узел – часть стебля, к которому прикрепляется лист или листья и почка или почки. Междоузлие – безлиственная часть стебля, которая обычно вытянута в длину. Узлы и междоузлия составляют такую часть побега как стебель. В составе побега есть боковые части или структурные (листья и почки). Онтогенез. В ходе полового размножения происходит слияние 2 гаплоидных гамет противоположного знака. Возникает диплоидная зигота. Из зиготы формируется зародыш. Зародыш – более или менее расчлененный миниатюрный спорофит – до начала быстрого роста находится некоторое время в состоянии относительного покоя. С началом активного роста зародыша начинается процесс органогенеза. Зародыш находится в семени и состоит из 2 взаимосвязанных осей – корневой и побеговой. Условной границей между ними можно считать место прикрепления I боковых листовых органов зародыша семядолей. Побеговая часть оси известна под названием почечки, или плюмулы. Она состоит из собственно осевой части – эпикотиля, 1 или нескольких листовых зачатков и апикальной меристемы побега. Зрелые семена при определенных условиях – достаточном количестве влаги, тепла, воздуха и света – прорастают. Прорастание начинается с увеличения всех частей зародыша за счет меристематической активности его клеток. Первым обычно появляется зародышевый корешок, укрепляющий молодое растение в почве. Семядоли в зависимости от типа прорастания ведут себя по-разному. Иногда они остаются в почве, и служат источником питательных веществ. В дркгих случаях семядоли выносятся на поверхность земли растущим стебельком и фотосинтезируют, выполняя функции первых листьев. В ходе дальнейшего развития на апикальной части первичного побега начинают закладываться новые листовые зачатки, в то время как ранее заложенные развертываются, а участки между ними разрастаются, образуя междоузлия. У проростков уже имеются основные вегетативные органы – корень и побег, которые продолжают нарастать за счет деятельности конусов нарастания.
Побег – самый изменчивый по внешнему облику орган растения. Основной тип побега зеленого растения – надземный ассимилирующий побег, несущий на оси нормально развитые зеленые листья. Однако наряду с фотосинтезом у побега бывают и иные функции: опорная, отложение запасных веществ, вегетативное размножение, образование цветков и соцветий. Одновременно со сменой функций могут меняться облик побега и характер его роста. Пример различного характера роста дают укороченные и удлиненные побеги. Первые характеризуются нормально развитыми междоузлиями и расставленными узлами, вторые – сближенными узлами. Укороченные побеги, в частности, имеют розеточные растения. На укороченных побегах нередко располагаются также соцветия.
В простейших случаях ветвления верхушка главной оси вильчато ветвится и дает начало 2 осям следующего порядка. Это так называемое верхушечное, или дихотомическое ветвление. Верхушечным ветвлением обладают многие многоклеточные водоросли, а также плауны. У остальных групп растений чаще встречается боковой тип ветвления. При этом на главной оси возникают боковые выросты, которые дают начало боковым осям. Следует выделить 2 типа бокового ветвления: моноподиальное и симподиальное. При моноподиальной системе ветвления каждая ось представляет собой моноподий, т.е. результат работы 1 верхушечной меристемы. Моноподиальное ветвление характерно для голосеменных и части травянистых покрытосемянных. Большинство покрытосеменных ветвится по симподиальному типу. Симподиальное ветвление формируется при повреждении верхушечной почки, при этом начинается усиленное развитие 1 или несколько боковых почек. Из них образуются боковые побеги, которые замещают побег, прекративший рост. Возкающая при этом ось является составной и сформирована из последовательной серии боковых побегов.
20.Лист – боковой структурный элемент побега. Морфология листа. Простые и сложные листья. Разнообразие строения листовых пластинок. Внутреннее строение листа. Функции листа.
Листорасположением, или филлотаксисом, называют порядок размещения листьев на оси побега. Закономерности размещения листьев связаны с деятельностью апекса побега и отражают его радиальную симметрию. Они имеют подчас довольно сложный характер. При характеристике листорасположения чаще всего ориентируются на количество листьев, отходящих от одного листового узла. Различают 3 основных типа листорасположения: спиральное, или очередное, - когда от каждого узла стебля отходит 1 лист (дуб, береза, злаки, зонтичные), супротивное – когда на каждом узле сидят друг против друга 2 листа (клен, сирень, губоцветные), и мутовчатое – когда каждый узел несет 3 и более листьев (олеандр, элодея). Листорасположение – наследственный признак. Иногда определенным типом листорасположения характеризуются целые семейства. В процессе роста побега листорасположение может меняться. Кроме того, черешки листьев могут изгибаться, и в силу этого положение пластинок также изменяется. При этом пластинки всех листьев в конечном итоге располагаются. Не затеняя друг друга, но образуют единую плоскость, где просветы между крупными листьями заполнены более мелкими листьями. Подобное явление, получившее название листовой мозаики, позволяет растению более плотно использовать падающий на него солнечный свет. Общая закономерность всех типов листорасположения – равное угловое расстояние между листьями, сидящими на одном узле или на последовательных узлах основной генетической спирали. Основной генетической спиралью, или парастихой, называют условную спиральную линию, которой можно соединить основания последовательных листьев. При очередном листорасположении на основной генетической спирали через 1 или несколько ее поворотов вокруг оси побега 1 лист оказывается более или менее строго под другим. Прямая, соединяющая листья, расположенные друг над другом, называется ортостихой. Участок спирали между 2 листьями на 1 ортостихе считается листовым циклом. Как число листьев в 1 цикле, так и число витков в нем характерно для каждого вида и имеет свое математическое выражение. Очередное листорасположение можно выразить дробью, где в числителе – число оборотов спирали в листовом цикле, а в знаменателе – число листьев в листовом цикле. Так, у злаков листорасположение выражается дробью ½; многие деревья: груша, слива, некоторые яблони имеют листорасположение, выражающееся дробью 2/5. Здесь листовой цикл имеет 2 оборота, в которых включены 5 листьев. На всем протяжении спирали при любом типе листорасположения листья сидят под строго определенным углом друг к другу, называемым углом расхождения.
Лист – это боковая часть побега, имеющая уплощённую форму и билатеральную симметрию. Лист выполняет 3 функции: фотосинтез; траспирация; газообмен. Лист- плоский орган, форма и размеры которого способствуют созданию максимальной фотосинтезирующей поверхности. лист двусторонний: верхняя и нижняя стороны. Верхнюю сторону называют брюшной или адаксиальной, а нижнюю - абаксиальной. Верхняя и нижняя стороны нередко различаются между собой по анатомическому строению, опушению, характеру жилкования. Размеры листа колеблются в пределах от 3 до 15 см. Взрослый лист расчленен на пластинку или несколько пластинок и черешок - узкую стеблевидную часть, соединяющую пластинку и узел побега. Нижняя часть листа, сочленная со стеблем называется основанием. Часто при основании есть прилистники. Пластинка- главная часть листа, осуществляющая основные функции. Черешок округлый или сплюснутый в поперечном сечении. Нередко черешок не развивается, тогда лист называется сидячим. Лист с черешком называется черешковым. Нередко основание листа разрастается и образует трубку, называемую листовым влагалищем. Листовое влагалище развивается при длительном сохранении интеркалярной меристемы листа.
Лист, имеющий одну пластинку, называется простым. В составе сложных листьев всегда несколько листовых пластинок, каждая из которых имеет свой черешок, а он крепится к общему черешку (рахис). В зависимости от расположения листочков различают перисто- и пальчатосложные листья. У перистосложных листочки располагаются двумя рядами по обе стороны рахиса. У пальчатосложный и тройчатосложных листьев рахиса нет, листочки отходят от верхушки черешка. По степени разветвления рахиса различают однократно, дважды и трижды перистосложные листья. Если рахис перистосложного листа завершается непарным листочков, лист является непарно- перистосложным, при отсутствии верхушечного листочка - парно- перистосложные.
Лист закладывается в основании апекса побега в виде бокового выступа- листового бугорка, затем превращается в листовой примордий. С этого момента начинается внутрипочечная фаза развития. Дальнейшее развитие листового примордия у разных типов листьев неодинаково. У части листьев в основании примордия закладываются прилистники. У простых цельнокрайних листьев примордий вытягивается и превращается в ось листа- дальнейшем срединную жилку, по бокам которой в результате краевого роста формируется пластинка. Во время роста примордия в нем дифференцируется проводящая система. Черешок развивается позднее других частей листа. С момента развертывания почки начинается внепочечная фаза. Поверхность листьев при этом увеличивается. У листьев двудольных это происходит за счет равномерного вставочного роста. Достигнув окончательных размеров, зеленые ассимилирующие листья живут различное время. У листопадных деревьев и кустарников, многолетних трав внепочечный период жизни 4-5 месяцев. Вечнозеленых растений субтропиков- от 2- 5 лет. Условие вечнозелености - развитие новых листьев тогда, когда еще не опали старые.
Пластинка листа или листочка может быть цельной или расчлененной. В зависимости от глубины расчленения выделяют лопастные, раздельные и рассеченные простые листья или листочки сложного листа Лопастный- расчлененный менее чем до половины ширины ( дуб). Раздельный - расчлененный глубже половины ширины ( одуванчик). Рассеченный- расчлененный до серединной жилки ( лютик). Расчленение осуществляется либо пальчато, либо перисто. Можно говорить о пальчато- и перистолопастных, пальчато- и перисторазделенных и пальчато- и перисторассеченных пластинках листьев. В первом случае выделяют лопасти, во втором - доли, в третьем- сегменты. Чаще встречаются однократно расчлененные листья,но у некоторых представителей семейств двудольных известны дважды, трижды и многократно расчлененные пластинки листа.
При характеристике листовой пластинки обращают внимание на ее форму, верхушку, основание и край. игольчатая (сосна), линейная (пшеница, ячмень), ланцетная (ива, олеандр, плаун), яйцевидная (граб, подорожник), округлая (осина), почковидная (копытень), сердцевидная (сирень), копьевидная (вьюнок), стреловидная (стрелолист), щитовидная (настурция). 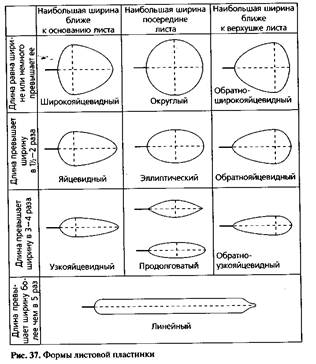
Если края листовой пластинки не имеют никаких выемок-лист цельнокрайним. Если выемки по краю листа неглубокие-лист цельным. Край листовой пластинки – пильчатый (острые углы).Край листовой пластинки – городчатый (округлые выступы).Край листовой пластинки – выемчатый (округлые выемки) 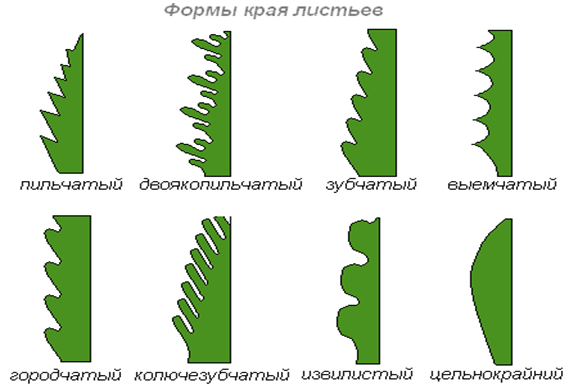
Жилкование- это система проводящих пучков и сопровождающих их тканей в листовых пластинках, посредством которых осуществляется транспорт веществ.В зависимости от особенностей прохождения продольных жилок выделяют параллельное и дуговидное жилкование. У двудольных известны два основных типа: перистое и пальчатое. Перистое: одна главная жилка, которая проходит от основания пластинки к ее верхушке. От главной жилки отходят боковые жилки первого порядка, от них- жилки второго порядка. Иногда боковые жилки первого порядка доходят до края пластинки и оканчиваются в лопастях, концах зубцов- это перистокраебежное жилкование ( береза, ольха). Перистопетлевидное: направляются к краю пластинки, но не достигают его, заворачивают дугой вперед, соединяясь со следующей боковой жилкой. Перистосетчатое: боковые жилки последовательно ветвятся, образуется густая сеть ( ивы, груши). Пальчатое: не имеется главная жилка, от места сочленения черешка и пластинки отходят несколько пальцевидно расходящихся крупных жилок первого порядка. Если доходят до края листовой пластинки- жилкование пальчатокраебежное, если вдоль края образуются петли- пальчатопетлевидное. При свободном ветвлении жилок второго и послед. порядка возникает пальчатосетчатое.
Основную массу листа, располагающуюся между верхней и нижней эпидермой, занимает хлоренхима, которая в листе получила особое название – мезофилл.
Цитологические особенности мезофилла:
1) клеточные стенки всегда тонкие (либо первичные, либо с незначительными вторичными изменениями),
2) наличие плазмодесм
3) наличие хлоропластов (их количество и размеры зависят от освещенности).
Форма клеток мезофилла может быть различной. У двудольных растений (особенно у светолюбивых) мезофилл дифференцирован на столбчатый и губчатый. Столбчатый мезофилл (палисадная паренхима) – слой плотно расположенных в 2-5 рядов слегка прозенхимных клеток. Эта ткань прилегает к верхней эпидерме, она содержит 75- 80% всех хлоропластов листа, поэтому здесь активно происходит световая фаза фотосинтеза.К нижней эпидерме прилегает губчатый мезофилл, он обычно включает 5-7 рядов округлых паренхимных леток, расположенных очень рыхло с большими межклетниками. Эти клетки также участвуют в фотосинтезе, но в большей мере осуществляют транспирацию. В состав губчатого мезофилла входят устьица. Благодаря развитой системе межклетников губчатого мезофилла СО2, поступающий через устьица внутрь листа, свободно распределяется по всей толще мезофилла, а выделяемый при фотосинтезе, по межклетникам поступает к устьицам и выделяется наружу. Внутренняя поверхность листа всегда во много раз превышает наружную поверхность листа. Так у ряда хвойных внутрь клеток мезофилла вдаются складки клеточной стенки и образуется так называемая складчатая паренхима.Дифференциация мезофилла на столбчатый и губчатый зависит от условий обитания. Проводящие ткани составляют основу жилок листа. Они состоят из первичных ксилемы и флоэмы, возникающих из прокамбия, объединены обычно в закрытые коллатеральные пучки. Пучки отделены от мезофилла листа крупными, часто лишенными хлорофилла обкладочными клетками. Механические ткани листа выполняют роль арматуры и противостоят его разрыву и раздавливанию. Обычно это склеренхимные волокна, сопровождающие все крупные проводящие пучки. В крупных жилках по краю листа нередко находится колленхима.
21.Корень, его функции, развитие, рост, ветвление. Понятие о корневых системах. Зоны корня, морфологические особенности, выполняемые функции.
Корень – вегетативный орган растения с радиальной симметрией и гетеротрофным питанием. Он способен к открытому росту до тех пор, пока сохраняется апикальная меристема. Морфологически корень отличается от побега тем, что на нём никогда не возникают листья, а апикальная меристема покрыта корневым чехликом.
Основные функции корня:
1. поглощение из почвы воды и минеральных веществ,
2. проведение воды и минеральных веществ в надземные органы, 3. заякоривание растений,
4. запасание основными тканями корня питательных веществ,
5. вегетативное размножение за счёт корневых отпрысков (малина, осина),
6. биосинтетическая – синтез фитогормонов, витаминов, алкалоидов, первичных АК.
Виды корней:
• главный корень, который всегда развивается из зародышевого корешка. У многих растений зародышевый корешок достаточно быстро отмирает, поэтому главный корень не развивается (злаки). У других видов главный корень выражен очень хорошо: одуванчик лекарственный, полынь горькая, почти все деревья.
• боковые корни образуются из перецикла на главном корне или боковых корнях более высокого порядка. При своём развитии боковые корни закладываются эндогенно, а затем прорывают первичную кору и выходят на поверхность главного или бокового корня. Благодаря боковым корням, происходит ветвление корневой системы. Главный корень – корень 1 пор., на нём развиваются боковые корни 2, 3 … порядка.
• придаточные корни никогда не развиваются на главном или боковых корнях. Они развиваются на стебле (смородина, традесканция), на листьях (узумбарская фиалка), на корневищах (ландыш майский, пырей ползучий, вороний глаз). Придаточные корни развиваются также при вегетативном размножении.
Корневая система — это совокупность всех корней растения.Типы корневых систем:
1. корневая система главного корня (стержневая корневая система). В этом случае главный корень всегда присутствует, и он имеет наибольшие размеры. Такая система характерна для большинства двудольных растений.
2. придаточная корневая система. В этом случае главный корень быстро отмирает, и на нижней части стебля (надземной) начинают активно развиваться придаточные корни.
• придаточная корневая система называется мочковатой, когда придаточные корни растут мочкой. Такая система характерна для однолетних злаков.
• у многолетних растений часто формируется корневище. Это подземное видоизменение стебля, значит от него отрастают только придаточные корни. Их совокупность образует еще одну разновидность придаточной корневой системы. Такая система характерна для ландыша, купены, клевера.
1. зона деления (несколько мм). Она отличается жёлтой окраской и представлена апексом корня, состоящим из мелких многогранных клеток – инициалей и их производных, эти клетки слабо дифференцированы, имеют густую цитоплазму и крупные ядра.
Снаружи зона деления прикрыта корневым чехликом. Он состоит из живых паренхимных клеток, возникающих у большинства однодольных растений из особой меристемы (калиптрогена), а у двудольных и голосеменных растений – из верхушечной меристемы кончика корня. Водные растения обычно лишены корневого чехлика.
Клетки чехлика живые, но поверхностные клетки периодически слущиваются и заменяются новыми, поступающими из зоны деления. Корневой чехлик выполняет несколько функций.
защищает апикальную меристему при продвижении корня через почву
клетки чехлика в процессе жизнедеятельности выделяют слизь, которая облегчает продвижение корня
в центральных клетках чехлика располагаются крупные амилопласты, и эти клетки способны воспринимать гравитацию.
Клетки корневого чехлика находятся в достаточно жёсткой среде, поэтому наружные клетки периодически отмирают и слущиваются. Новые клетки образуются из апикальной меристемы зоны деления.
2. зона растяжения, или роста (1-2см). Здесь клетки растягиваются, т. е. резко увеличиваются в размерах за счёт поступления воды и образования крупной центральной вакуоли. Наряду с процессами растяжения происходит и дифференцировка клеток, например, здесь начинает формироваться ризодерма. Зона роста выделяется светлой окраской и как бы прозрачна. Клетки этой зоны отличаются высоким тургором, что способствует раздвиганию частиц почвы.
3. зона всасывания, или зона корневых волосков (от нескольких мм до нескольких см) – все клетки полностью дифференцированы и закончили свой рост. Внутреннее строение этой зоны первичное, снаружи она покрыта эпиблемой (ризодермой), которая активно поглощает воду и минеральные соли. В составе ризодермы находятся трихобласты и атрихобласты, которые увеличивают поверхность всасывания примерно в 18 раз.
4. зона проведения – наиболее протяжённая зона. У корней однодольных растений и в молодой части древесных растений зона проведения покрыта экзодермой. Более старая часть корня покрыта перидермой, (у древесных растений старые части корня могут быть покрыты пробкой). У двудольных растений зона проведения имеет вторичное анатомическое строение. Основная функция – вертикальное проведение из почвы в побег водного раствора, а также проведение органических веществ в зоны растяжения и роста.
22.Первичное и вторичное анатомическое строение корня, гистологические особенности, использование характерных черт анатомического строения корней как диагностических признаков.
В первичном строении корня отмечают три анатомо-топографических зоны: покровы, первичная кора, центральный цилиндр (стель).
Для первичного строения корня характерно хорошее развитие первичной коры. В центре стели располагаются прочные элементы: либо первичная ксилема, либо склеренхима.
Покровы представлены ризодермой, в состав которой входят трихобласты с корневыми волосками, а также атрихобласты.
Первичная кора, расположенная под покровами хорошо выражена, в её составе выделяют три ткани:
• экзодерма – 1 ряд паренхимных плотно (без межклетников) расположенных клеток. В зоне проведения, после того как клетки ризодермы отомрут и слущатся, зкзодерма будет выполнять функцию покровов.
• мезодерма – несколько рядов рыхло расположенных паренхимных клеток, в которых накапливаются запасные питательные вещества. Мезодерма (а также экзодерма) обеспечивают радиальный транспорт водного раствора от ризодермы к центральному цилиндру.
• эндодерма – внутренний слой коры корня, граничащий с центральным цилиндром. Она представлена одним слоем толстостенных клеток, большая часть которых имеет пояски Каспари. Пояски Каспари – это формирующаяся вторичная клеточная стенка. Она пропитывается суберином, сначала суберин расположен пятнами, а потом пятна увеличиваются в размерах и формируют пояски. Клетки с поясками Каспари не способны проводить воду из мезодермы в центральный цилиндр, т.к. их протопласт отмирает. Зрелая эндодерма на 75-80% состоит из таких клеток. Остальные клетки эндодермы живые с тонкими неодревесневшими стенками. Это пропускные клетки: через них осуществляется связь первичной коры с осевым цилиндром.
В центральном цилиндре самая наружная ткань представлена перециклом. Это первичная меристема, состоящая из 2-3 слоёв клеток. Большую часть стеля занимает радиальный проводящий пучок (у двудольных может быть 1- 5 лучей, у однодольных – от шести и более). В самом центре пучка расположена первичная ксилема в форме 4-х лучевой звезды, а между лучами ксилемы находятся участки первичной флоэмы и некоторое количество основной паренхимы.
Такой тип строения органа, где водопроводящие и механические элементы располагаются в самом центре, обеспечивает достаточную прочность корня на разрыв. В состав центрального цилиндра также входят клетки основной паренхимы.
У однодольных растений первичное строение корня сохраняется на протяжении всей жизни. У двудольных растений и голосеменных достаточно быстро закладывается камбий и формируется вторичное строение корня. Его легко наблюдать на уровне зоны проведения. Особенностью развития камбия в корне является то, что он закладывается в виде лОпастного кольца между участками первичной ксилемы и флоэмы. Частично камбий образуется из перецикла, а частично из уже дифференцированных клеток основной паренхимы, расположенной между первичной и вторичной флоэмой. Деятельность камбия приводит к тому, что кнаружи откладывается вторичная флоэма, а внутрь вторичная ксилема. В итоге формируется вторичный коллатеральный проводящий пучок, в котором вторичная ксилема и вторичная флоэма расположены друг против друга. Пучки разделены вторичной запасающей паренхимой. Таким образом, камбий обеспечивает вторичное утолщение корня, которое со временем может стать значительным.В результате развития вторичных тканей первичная ксилема, которая представлена радиальным пучком, полностью сохраняется в центре корня. Первичная флоэма отодвигается вторичными проводящими тканями к переферии и постепенно сдавливается и разрушается (часто полностью). Судьба первичной коры может быть разной. Очень часто под напором боковых корней первичная кора разрывается на фрагменты, а затем полностью слущивается. В этом случае из остатков первичной коры может возникнуть феллоген. Феллоген откладывает наружу клетки, которые вскоре отмирают и образуют пробку. Оказавшиеся снаружи от пробки живые клетки первичной коры также отмирают, после чего корень приобретает буро-коричневую окраску. Такое происходит у корней всех многолетних растений (на второй год). У молодых корней первого года жизни первичная кора в зоне проведения может сохраниться, и её наружные клетки будут выполнять функцию покровов.
|
из
5.00
|

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы