 |
Главная |


Ретикулярная формация и гиго галамус. Висцерально-эндокринные функции
|
из
5.00
|
Одной из важнейших сторон деятельности ретикулярной формации является ее прямое и опосредованное участие в регуляции ряда жизненно важных функций, как например, дыхания и кровообращения. Не менее важно влияние и на целый ряд других функций: висцеральных, эндокринных и защитных.
Начнем с функции внешнего дыхания, как наиболее моторной из всех висцеральных функций. Регуляция внешнего дыхания или вентиляция легких по существу сводится к регуляции моторных функций дыхательных мышц, к которым относятся в первую очередь межреберные мышцы и диафрагма. Мотонейроны шейного и грудного отделов спинного мозга, иннервирующие диафрагму и межреберные мышцы, получают управляющие импульсы от бульбарного дыхательного центра, расположенного в ретикулярной формации, на дне IV желудочка, в каудальной части продолговатого мозга (рис. 19). Понятие «дыхательный центр», как системы регуляции внешнего дыхания, достаточно сложное, так как объединяет в себе различные структуры мозга, расположенные начиная от коры больших полушарий и кончая спинным мозгом. Однако чаще всего говоря о дыхательном центре, имеют в виду ретикулярную формацию варолиева моста (пневмотаксический центр) и бульбарного отдела ствола мозга (дыхательный моторный центр), •которую впервые подробно исследовал Н. А. Миславский еще в 1885 г.
Несмотря на продолжающиеся споры о локализации и нейронном строении ретикулярных структур, ответственных за вдох и выдох, тем не менее все авторы согласны, что эти два важнейших отдела бульбарного дыхательного центра располагаются в пределах субвентрикулярного серого вещества (рис. 19), в гигантоклеточном ретикулярном ядре, захватывая частично вентральные и латеральные ретикулярные ядра (Pitts, 1946; Brodal,..J957; Torvik, Brcda'l, 1957; Vvaiig, Ngai, 1963) -
Мы не будем останавливаться на вопросах морфо-ана-томии бульбарного дыхательного центра, на спорных и нерешенных проблемах, так как нас интересует в данном случае тот факт, что дыхательный центр, принадлежащий ретикулярной формации, является главным автоматическим регулятором дыхательных движений. Именно из этой части-ретикулярной формации начинаются нисходящие ретику-ло-спинальные волокна к мотонейронам дыхательных мышц шейных и грудных отделов спинного мозга.
Ретикуло-спинальные влияния на другие скелетные мышцы (главным образом локомоторные) являются лишь

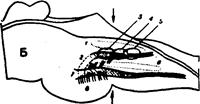
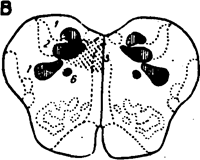
Рис. 19. Дыхательный центр.
А—на горизонтальном плане видны контуры моторных и сенсорных ядер Х и V нервов и зоны вдоха (справа) и выдоха (слева).
Б—на сагиттальном плане видны те же ядра, что и на рисунке <А», а также волокна Х и V нервов.
На солитарном тракте обозначены участки, где сконцентрированы волокна к зонам выдоха (слева), вдоха (справа) и деп-рессорный зоне (посередине) сосудодвигательно-го центра.
В — на фронтальном плане видны те же ядра, что и на рисунке «А», а также зона выдоха.
Условные обозначения:
1 — дорзальное моторное ядро Х нерва, 2—соли-тарный тракт и его ядро;
3—зона выдоха, 4—деп-рвсодрная зона; 5 — зона вдоха; 6— обоюдное ядро Х нерва; 7 — спинальный тракт и ядро V нерва;
8—'афферентные волокна, идущие к тригеминально-му и солитарному ядрам.
тонизирующими, но не пусковыми, как уже подчеркивалось, то есть ретикулярная формация не диктует движение, а лишь управляет его уровнем, приспосабливая мышечный тонус к реальному характеру мышечной нагрузки. Для дыхательных мышц ретикулярная формация является как пусковой, так и тонизирующей системой, то есть вызывает само движение и управляет его характером. Поэтому сильное синхронное возбуждение ретикулярной формации, например при болевой стимуляции, переключаемое ьа дыхательный центр, может рефлекторно запустить остановившееся дыхание, даже при выключенном сознании.
Мы не будем разбирать также огромную литературу по физиологии дыхательного центра и отдельных его нейронов, так как нас интересует главным образом рефлекторный механизм регуляции дыхательных движений и других функций аппарата дыхания. В связи с этим важно отметить, что бульбарный дыхательный центр у всех позвоночных вплоть до человека располагается рядом с сенсорными и моторными ядрами блуждающего нерва (рис. 19): солитар-ным ядром, дорзальным ядром и обоюдным яд^ом (п. ат-biguus), так как блуждающий нерв является основным коллектором обгспечения рефлекторных реакций различных органов, участвующих в обеспечении стандартной внутренней среды, а дыхание — одно из важнейших функций обеспечения гомеостаза (Е. К. Сепп, 1959; Oberholzer et al., 1960). Хотя дыхательный центр обладает собственным автоматизмом (Salmoiraghi, von Baumgarten, 1961), который может осуществляться даже при выключенном сознании (во сне), однако на его работу влияет целый ряд экстраретикулярных факторов, среди которых важнейшими являются афферентные сигналы, передаваемые волокнами блуждающего нерва (X) огрецепторов растяжения легких (рефлекс Геринга-Брейера); сигналы от рецепторов дыхательных путей, передаваемые волокнами V, VII, IX, Х нервов;
сигналы от хеморецепторов каротидного клубочка по веточкам языко-глоточного нерва (IX), при уменьшении парциального давления Оа в крови или увеличении парциального давления СОа. Помимо рефлекторных реакций важнейшим фактором воздействия непосредственно на нейроны дыхательного центра является состав крови, особенно избыток С02. Не касаясь химических факторов регуляции дыхания, отметим, что вся афферентная рефлекторная регуляция дыхания осуществляется при участии нервных волокон вагуса и всех других аффереьтов, участвующих в иннервации ушной раковины. В этом нет ничего странного и неожиданного, если вспомнить эволюцию ушной раковины и ее нервного аппарата. Дыхательный центр или, точнее, ретикулярная формация, в состав которой входит дыхательный центр, располагается как раз в непосредственной близости от солитарного тракта и его ядра (рис. 19), которое является сенсорным центром вагуса (X) и остальных нервов ушной раковины (V, VII, IX). Наличие коллатералей в ретикулярную формацию, особенно от афферентов вагуса и вторичных волокон солитарного и спинального тригеми-нального ядер, обеспечивает рефлекторную активацию окружающей ретикулярной формации и расположенного в ней дыхательного центра, как и других рефлекторных центров ствола мозга.
На рисунке 19 видны тесные связи солитарного тракта и его ядра (посредством коллатералей вторичных афферентов) с экспираторной и инспираторной областям дыхательного центра, а также с депрессорным отделом сосудодви-гательного центра.
Ниже солитарного тракта расположен спинальный (нисходящий) тракт и ядро тройничного нерва и видно, как часть волокон Х нерва вместе с волокнами V нерва заходят и в это ядро, и в солитарное ядро.
Рефлекторные реакции на дыхательный центр (на функцию дыхания) имеют место при стимуляции любых афферентных систем (В. Н. Черниговский, 1960), но при стимуляции афферентов блуждающего нерва и других афферентов солитарного тракта эти рефлексы особенно выражены. Достаточно эффективны болевые стимулы для рефлекторного воздействия на дыхательный центр. Рефлекторные реакции в дыхательной системе, связанные с ретикулярными нисходящими эффектами, распространяются ье только на дыхательные мотонейроны спиьального уровня, но и на ряд других функций, участвующих в интеграции дыхательного акта, который, конечно, не состоит только из сокращения межреберных мышц и диафрагмы. В регуляции функции дыхания важное значение имеет состояние тонуса гладкой мускулатуры бронхов, альвеолярное кровообращение и артериальное давление в малом круге кровообращения, проницаемость альвеолярных и капиллярных стенок для газов крови и др. Все это также контролируется и регулируется ретикулярной формацией ствола и запускается, по-видимому, через рефлексы дыхательного центра.
Таким образом, афферентные системы блуждающегоX,тройничного (V) и языко-глоточного (IX) нервов, которые участвуют в иннервации ушной раковины, оказывают мощный рефлекторный эффект на функции дыхания. Благодаря такой физиологической закономерности аурнкулярные рефлексы оказываются способными нормализовать нарушение функции в системе органов дыхания. Например, при лечении бронхиальной астмы важнейшими условиями нормализации функции дыхания являются снятие гипертонуса гладкой мускулатуры бронхов и подавление гиперсекреции слизистой бронхов, что достигается рефлекторно, через воздействие на соответствующие центры ретикулярной формации. Восходящие эффекты ретикулярной формации, передаваемые в гипоталамус, приводят к активации системы гипофиз-кора надпочечников, благодаря чему постепенно нормализуется также гормональная функция, которая играет существунную роль в патогенезе бронхиальной астмы.
Итак, тесная связь аурикулярных афферентов с ретикулярной формацией ствола мозга через два самых мощных афферентных центра, каковыми являются тригеминальные ядра и солитарное ядро, которое формировалось и совершенствовалось в процессе эволюции, является физиологическим фундаментом рефлекторных реакций различных ретикулярных структур, в том числе дыхательного центра.
Рядом с дыхательным центром располагается другой жизненно важный ретикулярный центр ствола мозга — сосудодвигательный (рис. 20), который регулирует частоту сердечных сокращений и сосудистый тонус (Uvnas, 1960;
Ochs, 1965; Ganong, 1969). Сосудодвигательный центр образован двумя важнейшими областями: прессорной, которая усиливает тонус гладкой мускулатуры сосудов и повышает артериальное давление. Более каудально от прессорной области расположена область, которая снижает сосудистый тонус и способствует понижению артериального давления. К сосудодвигательному центру подходят волокна от гипоталамуса, раздражение которого вызывает учащение сердцебиений и повышение артериального давления.
Эти влияния передаются на сердце и сосуды через симпатические нервы (рис. 21). При раздражении медиального гипоталамуса можно получить и депрессорные эффекты на сосуды, например, расширение сосудов скелетных мышц. Прессорно-депрессорные эффекты осуществляются также с областей, расположенных в лимбической системе (Ganong, 1969).
Наконец, сердечно-сосудистые реакции можно вызвать и при раздражении коры (моторной области, лобной области, височной области, поясной извилины лимбического мозга). Корковые влияния на сосудодвигательный центр .
Рис. 20. Сосудодвигатель-ный центр.

А—на горизонтальном плане видны заштрихованные контуры прессор-ной зоны (1), депрессор-ной зсны (2), зоны тормо-2 жения сердечной деятельности (3), образованной • нейронами дорзального моторного ядра Х нерва, зоны рефлекторных эффектов сосудистых хемо-рецепторов и болевых сигналов от сомато-вис-церальных афферентов (4), зона рефлекторных эффектов каротидных и аортальных барорецепто-ров (5).
Б — на фронтальном плане видны заштрихованные зоны прессорной (1) и депрессорной (2) зон, а также дорзальное моторное ядро Х нерва (6), солитарный тракт и его ядро (7), обоюдное ядро . „ Х нерва (8), клиновидное
ядро дорзальных столбов спинного мозга (в), • 'спинальный тракт и
ядро V нерва (10).
носят сугубо рефлекторный характер и не являются произвольными, как это имеет место при корковых эффектах на дыхательный центр. Человек может произвольно регулировать характер дыхания и даже остановить его (на короткое время, пока концентрация С02 в крови достигнет критического уровня), но произвольно менять ритм сердечных сокращений или тонус сосудов в обычных условиях невозможно (в исключительных случаях это удается отдельным йогам, после упорной и длительной тренировки). Однако наличие кортикофугальных эффектов на ретикулярную формацию потенциально может способствовать тому, чтобы путем контролируемого обучения на основе
Рис. 21. Схематические ^. КОРА «вязи сосудодвигательно- .^-^1^ го центра. //^~^\

Сокращения и условные обозначения: Гипо- \ тал.— гипоталамус; Ср. ^ мозг. — средний мэзг, \ Пр. мозг.—продолговатый ГИПОТАА. \ мозг; IX, X— черепномоз-говые афферентные нервы;
стрелками указано направление нервных сигналов.
1 — прессорная зона и 2 — депрессорная зона, откуда начинаются рети-«уло-спинальные волокна на симпатические элементы спинного мозга; 3 — тормозящая зона сердца <дорзальное моторное ядро Х нерва); 4 — сердечные ветви Х нерва из дорзального моторного ядра; 5 — прессорные и 6 — депрессорные рети-куло-спинальные волокна «а симпатические элементы спинного мозга; 7 — симпатические узлы и цепочка; 8 — симпатические нервы сердца; 9 — симпатические нервы сосудов.
обратной связи (biofeedback) обучить человека произвольно контролировать уровень артериального давления. Рефлекторный характер кортикофугальных эффектов на со-судодвигательный центр связан главным образом с корковым запуском двигательной мышечной активности, для обеспечения которой требуется изменение мышечного кровообращения. Хотя дыхание также должно приспосабливаться к интенсификации мышечных движений, однако это лучше достигается путем воздействия на дыхательный центр газами крови, а для сердечно-сосудистой системы программа действий задается корой, одновременно с запуском движений. Учитывая эту физиологическую особенность при рефлексотерапии мышечных нарушений, особенно связанных с кровообращением, необходимо обязательно принимать меры для усиления мышечного кровотока. Здесь, как и в случае воздействия на мышечный тонус, целесообразно стимулировать также аурикулярные точки «коры» и «гипоталамуса» (рис. 39).
Дорзальное моторное ядро блуждающего нерва является одновременно центром подавления сердечной деятельности. В это ядро идут сигналы по афферентам барорецеп-торов сердца и крупных сосудов; поэтому при повышении артериального давления рефлекторно замедляется сердечная деятельность через сердечные (холинергические) ветви эффекторной части блуждающего нерва, которые берут начало в этом ядре (рис. 21). Усиление сердечной деятельности осуществляется симпатическими (адренергическими) нервами сердца через сссудодвигательный (сердечно-сосудистый) центр благодаря тому, что рефлекторно усиливается тонус этой области ретикулярной формации и одновременно подавляется активность нейронов, замедляющих сердечную деятельность в дорзальном моторном ядре Х нерва. На нейроны этого ядра оказывает рефлекторное влияние не только афференты вагуса, но и другие афферен-ты солитарного тракта, среди которых представлены нервы ушной раковины.
Следовательно, аурикулярные воздействия, особенно на афференты Х и IX нервов, должны вызывать выраженные рефлексы на сердечно-сосудистую систему. Эти рефлексы должны быть особенно интенсивными при стимуляции внутреннего края противозавитка (со стороны конхи), где представлены, помимо указанных, также терминали тройничного нерва. Эта зона расположена напротив корня завитка или «чудесной точки» (Nogier, 1972) рядом с представительством седьмого шейного и первого-второго грудных позвонков (рис. 22). Nogier располагает точку сердечного ритма в полости конхи, у самого входа в наружный слуховой проход, одновременно указывая также точку сердца. Эти обе точки достаточно эффективны и во время их стимуляции наблюдается кратковременное (3—5 секунд) усиление амплитуды пульсовой волны (сердечного выброса) на лучевой артерии, что послужило одним из поводов говорить о существовании аурикуло-кардиального рефлекса. Однако зона, указанная нами, еще более эффективна и при ее стимуляции наблюдается более стойкий и 112 выраженный эффект учащения ритма сердечных сокращений и усиление сердечного выбрсса (рис. 22).

а) б) в)
Рис. 22. Специальные точки и зоны ушной раковины:
а — для реанимации;
б—для мышечного тонуса: черные — в области иннервации V нерва; белые— в области иннервации нервов Cg—Сд;
в — для сердечного-сосудкстых рефлексов: 1 —t нулевая точка» 2—«чудесная точка»; 3 — точка сердца; 4— точка сердечного ритма.
Для того, чтобы не впасть в ошибку, следует учесть, что кардио'васкулярный рефлекс имеет место всегда при неожиданном раздражении (даже слабом), при болевом раздражении, при сильном волнении, страхе и т. п. Системы дыхания и кровообращения, как наиболее лабильные,. всегда реагируют на любий синхронный афферентный залп по соматс-висцеральным волокнам или коллатералям, на неожиданные экстрарецептивные сигналы (зрительные,слу-ховые, обонятельные), на эмоционально значимые стимуляции (Э, А. Костандов, 1977). Дыхательный и сердечнососудистый рефлекс всегда является компонентом ориентировочной реакции и носит уже кортикофугальный характер.
Поэтому терапевтически значимыми рефлексами на сердечно-сосудистую и дыхательную системы при аурикуляр-ной рефлексотерапии (как и при корпоральной) следует считать не кратковременный (несколько секунд) эффект (аурикуло-кардиадьный рефлекс), который будет иметь место почти всегда, как неспецифический компонент любой и рефлекторной реакции. Терапевтически значимыми являются длительные, функциональные изменения, которые измеряются не секундами, а минутами и часами, действующие в период продолжающегося воздействия на ушную раковину и после него.
В числе важнейших ретикулярных рефлексов, связанных с функциями дыхания, следует назвать такие сложно-координированные акты, как чихание, кашель, зевота. Первые два рефлекса возникают при раздражении слизистых дыхательных путей и носят защитный характер, обеспечивая удаление различных посторонних агентов. Афферентные сигналы передаются по волокнам V, IX и Х нервов. При чихании (например, при попадании паров аммиака в носоглотку) афферентные сигналы передаются по волокнам V нерва, вызывают остановку дыхания в состоянии вдоха и сужение бронхов, после чего воздух под давлением выбрасывается наружу. Остановка дыхания и суже-яие бронхов при чихании еще раз свидетельствуют, что возбуждение тройничного нерва оказывает влияние не только на дыхательный центр, но и на моторное ядро блуждающего нерва. Это обстоятельство свидетельствует также, что рефлекторное воздействие на тонус гладкой мускулатуры бронхов при стимуляции афферентов не только X, •но и V нерва физиологически вполне детерминировано, что в свою очередь объясняет эффективность аурикулярной рефлексотерапии бронхиальной астмы.
Наконец, существование рефлекторного эффекта с аф-<рерентов V нерва на двигательное ядро Х нерва и на близлежащую ретикулярную формацию указывает на то, что при стимуляции ушной раковины и возбуждении чувствительных окончаний V нерва и других нервов уха неизбежны различные рефлексУна двигательные ядра Х нерва, на ретикулярные центры и связанные с ними функции.
К числу таких функций относится также акт рвоты, который осуществляется сложно-координированными двигательными (сократительными) реакциями мышц кишечника, желудка, брюшной стенки, диафрагмы, глотки.
Афферентные сигналы поступают по волокнам Х и IX нервов в солитарное ядро от слизистой желудочно-кишеч-ного тракта, от ротовой полости, глотки и языка и переключаются на дорзальное моторное ядро Х нерва, которое расположено в окружении дыхательного и сосудодвигатель-ного центров. Коллатерали волокон дорзального ядра активируют ретикулярную формацию, в частности дыхательный центр, в связи с тем, что двигательный компонент рвотного акта обеспечивается не только сокращениями стенок желудочно-кишечного тракта, но также дыхат-льных мышц и мышц брюшного пресса (Eliasson, 1960).
Таким образом, как и в случае с рефлексом «к&шель-чихание», в запуске рвотного рефлекс участвуют те же афференты, которые иннервируют ушную раковину, что является одним из аргументов, объясняющих, почему воздействие на ушную раковину вызывает моторные реакции желудочно-кишечного тракта, межреберных, брюшных и диафрагмальных мышц. Столь тесная связь афферентов ушной раковины, в частности Х и IX нервов, с указанными выше функциями объясняется тем, что в эволюции эти нервы имели прямое отношение к иннервации ротовой полости, глотки и жаберного аппарата и участвовали в рефлекторной защите этого органа от попадания инород-них тел и непригодной (в химическом отношении) для дыхания воды. Эта реакция у рыб является чем-то средним между кашлем-чиханием и рвотой млекопитающих (удаление инородних тел из дыхательных путей- и извержение-наружу вредного для организма содержимого).
Если в области дна IV желудочка, в бульбарной части ствола, расположены жизненно важные центры дыхания » кровообращения, то в области дна III желудочка, в диэнце-фальной части мозга, расположена группа специализированных ядер гипоталамуса, которые имеют прямое отношение к регуляции не менее важных функций внутренних органов и внутренней среды организма.
Велико также значение гипоталамо-гипофизарных вза* имоотношений в регуляции гормональных функций. Мы будем касаться этих вопросов лишь в той мере, которая необходима для понимания механизмов аурикулярной рефлексотерапии, тем более, что они подробно представлены в литературе (Я. Сентаготаи, Б. Флерко, Б. Меш, Б. Ха-лас, 1965; Harris, 1960; in^ram, 1960; Haymaker, Anderson, Nauta, 1969; Curtis, Jacobson, Marcus, 1972).
Мы уже знаем, что между гипоталамусом и ретикулярной формацией существуют тесные взаимосвязи и что такие же связи существуют у гипоталамуса с лимбической системой, таламическими ядрами и лобными отделами коры мозга. В предыдущих разделах, описывая те или иные функциональные механизмы, например, ретикулярной формации, мы не оговаривали важнейшее условие, что в целостном мозге, не отравленном наркотическими и другими психотронными препаратами, любые рефлекторные реакции осуществляются не локальными изолированными структурами мозга, а целой системой взаимосвязанных структур, находящихся друг с другом в сложной иерархии подчинения. Так, например, целый ряд ретикулярных рефлексов можно подавить при помощи коркового торможения. Кровообращение регулируется не одними автоматическими рефлексами ствола, но всеми надстволовыми структурами мозга:
гипоталамусом, лимбическим мозгом, корой мозга. Однако основные связи и механизмы реализации сосудодвига-
•тельного рефлекса представлены в ретикулярной формации
•ствола, поэтому при описании таких механизмов с целью их упрощения и большей доходчивости можно пренебречь многоступенчатыми связями, но всегда помнить, что они
•есть. Такой принцип, несколько облегчающий изложение
•сложного материала, мы будем использовать и дальше, в частности при рассмотрении гипоталамических механизмов.
В латеральном гипоталамусе находится центр «голода», а в вентро-медиальном — центр «насыщения» (Brobek, 1960;
Anand, 1961). Между этими центрами существуют реципрок-ные взаимоотношения, благодаря чему при пищевом насыщении постепенно усиливается активность центра «насыщения» и одновременно подавляется активность центра «голода» (Eliasson, 1960). При этом активный центр «насыщения» при помощи гипоталамо-ретикулярных связей подавляет двигательную активность, снижает тонус ретикулярной формации и клонит ко сну (Ochs, 1965). Напротив,
•возбужденный центр «голода» усиливает двигательную реакцию, повышает сенсорные функции, настраивая организм на активный поиск пищи. Организм человека, хотя
•также подчинен этим закономерностям, однако корковый
•контроль может вносить свои коррективы. Тем не менее голодный человек также стремится утолить голод. Если .искусственно изменить возбудимость этих центров, например, при стимуляции гипоталамических ядер вживленными электродами, то резко меняется пищевое поведение. Например, при возбуждении центра «насыщения» или удалении центра «голода» развивается гипофагия или даже афа-гая. Следовательно, при рефлекторном возбуждении вентро-медиального гипоталамуса (центр «насыщения») можно резко снизить аппетит и таким образом вылечить от гипер-^)агии. Выход коллатералей аурикулярных афферентов через центральное серое вещество ствола в гипоталамус объясняет возможность аурикулотерапии ряда функций, регулируемых гипоталамусом, в том числе пищевой мотивации (К. В. Судаков, 1971).
Гипоталамус является основным центром автоматического регулирования всех внутренних процессов организма и в первую очередь внутренней секреции, деятельности органов брюшной полости и малого таза, а \акже сердечнососудистой системы. Хотя гипоталамус входит в состав висцерального мозга и контролируется его корковыми структурами, тем не менее гипоталамус в значительной степени является самостоятельным рефлекторным центром •(Papez. 1958). Благодаря этому целый ряд висцералыю-эндокринных функций могут успешно управляться рефлекторными реакциями гипоталамуса (Я. Сентаготаи, Б. Флерко, Б. Меш, Б. Халас, 1965).
Помимо регуляции количества потребляемой пищи, гипоталамус участвует также в регуляции уровня метаГо-лизма, согласуя его с потребностями организма, его энергозатратами на протяжении длительного времени, о чем свидетельствует относительное постоянство веса. Избыточный вес (ожирение) связан не только с потреблением большого количества пищи, но и с извращением метаболизма. Участие гипоталамуса в регулировании обоих факторов является основанием для того, чтобы нормализовать эти нарушения методами рефлекторного воздействия, особенно через афференты ушной раковины, проекции которых, как известно, активируют мезенцефалическую ретикулярную формацию и гипоталамус.
Важнейшей функцией гипоталамуса является способность регулировать водно-солевой баланс. Супраоптические и латеральные ядра гипоталамуса (рис. 23) имеют прямое отношение к регулированию содержания воды в организме. При искусственном разрушении супраоптического ядра <эксперимент) или патологических явлениях в области этого ядра и его связей с задн и долей гипофиза возникает несахарный диабет, который выражается в полидипсии (до 40 л воды в сутки) и полиурии. В заднем гипофизе вырабатывается вазопрессин или антидиуретический гормон <АДГ), который действует на почки и стимулирует обратное всасывание воды в почечных канальцах, то есть регулирует процесс концентрирования мочи. Уровень синтеза АДГ регулируется гипоталамусом «благодаря наличию в нем специальных осморецептивных нейронов, которые реагируют на степень сгущения крови и передают соответствующие сигналы в гипофиз (Sundsten, Sawyer, 1961).

Рис. 23. Гипоталамо-гипофизарные взаимоотношения.
Условные обозначения: Hth — гипоталамус, HVp — гипофиз; А — аденогипофиз; N — нейрогипофиз; 1 — мамилярное ядро; 2 — заднее ядро; 3 — дорзо-м°диальное ядро; 4 — вентро-медиальное ядро; 5 — па-авентрикулярное ядро; б—переднее ядро; 7 — супраопткческое ядро; 8 — преоптическая область; 9 — хиазма оптика; 10 — портальная капиллярная система; 11 — гипоталамо-гипофизарный тракт.
Следовательно, регулирование синтеза АДГ в системе гипоталамус-гипофиз для управления реабсорбционной функцией почек может быть достигнуто либо за счет .рефлекторного стимулирования супраоптической области гипоталамуса при воздействии на соответствующие точки ушной раковины, либо за счет рефлекторного изменения возбудимости осморецепторов или, точнее, осмочувствитель-ных нейронов гипоталамуса. На самом деле оба механизма работают, видимо, одновременно.
Контроль за содержанием воды в организме осуществляется не только путем воздействия на функции почек, но также путем регуляции потребления воды. В латеральном гипоталамусе (рис. 23) имеется центр жажды, подобно центру голода, при стимуляции которого потребление воды увеличивается, а при торможении — сокращается (Anand, 1961). Следовательно, используя этот механизм, можно. научиться рефлекторно регулировать жажду; можно добиться, чтобы в соответствующих условиях человек меньше ощущал потребность в воде, чем это бывает обычно, можно подавить чувство голода и жажды до определенных пределов (не угрожающих жизненным нормам).
Наконец, следует отметить, что водный баланс связан с потреблением солей, особенно электролитов Na+ и К*. уровень которых в организме регулируется альдостероном— гормоном коры надпочечников. В отличие от других гормонов коры надпочечников, синтез которых теснейшим образом связан с гипофизом, альдостерон, принадлежащий к минералокортикоидам, регулируется гормоном, который образуется в мезенцефалической ретикулярной формации <Farrell, 1958; К. Лишшак, Э. Эндрёци, 1967).
Учитывая большие потенциальные возможности аури-кулярных рефлексов на функции гипоталамуса и ретикулярной формации, в том числе мезенцефалической, можно <5ыть уверенным в реальной возможности рефлекторно влиять на водносолевой баланс организма: на чувство жажды и потребность воды, на функцию почек, усвоение и удержание в организме ионов Na*и К*.
Следовательно, при тех заболеваниях, при которых помимо основных симптомов имеются признаки нарушения метаболизма и водно-солевого баланса, необходимо применять стимуляцию таких точек ушной раковины, которые вызывают рефлекторные реакции гипоталамических механизмов. К ним относятся не только точки, расположенные в области конхи, но также в трехсторонней выемке, в ко зелковой и предкозелковой половине мочки уха. Проблема идентификации «гипоталамических» точек ушной раковины имеет огромное практическое значение и поэтому ее следует разрабатывать как в условиях экспериментов на животных. так и путем тщательных клинических наблюдений. Точки, которые рекомендованы в настоящее время (об этом см. ниже), недостаточно обоснованы и имеют ограниченное применение.
Так как в регулировании температуры тела большую роль играет сосудистая система, например, для теплоотдачи (Strom, 1960) путем расширения поверхностных крове-ностных сосудов и усиления конвекции и излучения (Hardy, 1961) нет ничего странного в •юм, что центр терморегуляции располагается в гипоталамусе, причем, как и в предыдущих случаях, имеются реципрокно работающие системы — центр теплоотдачи в переднем гипоталамусе и центр теплообразования в заднем гипоталамусе.
Гипоталамус «измеряет» температуру крови при помощи специальных термочувствительных элементов — нейронов и включает соответствующие механизмы регуляции. При охлаждении организма сенсорные импульсы от терморецепторов кожи действуют на гипоталамус, который включает механизмы стимулирования образования тепла (дрожь)" и уменьш 'ния теплоотдачи (сужение поверхностных сосудов и пилоэрекция). Повышение температуры при инфекционных заболеваниях связано с бактериальными эндотоксинами, которые обладают пирогенным эффектом и действуют, по-видимому, на гипоталамический центр теплообразования, усиливая его функцию. Следовательно, подавляя функцию этого центра апирогенными веществами (фарма-котерапия) или рефлекторным путем, можно снизить повышенную температуру.
Хотя «измерение» температуры крови осуществляется непосредственно нейронами гипоталамуса, термочувствительная информация передается по волокнам спино-тала-мического тракта или его тригеминального гомолога. При этом часть волокон или их коллатерали, передающие термические и ноцицептивные сигналы, проходят в мезен-цефалическом отделе в центральное серое вещество и далее в гипоталамус и таламус. В связи с этим есть все основания утверждать, что применяемые методы рефлекторного обезболивания, реализуемые в значительной степени в мезен-цефалическом сером веществе, в интраламинарных ядрах таламуса и в гипоталамусе (Fairman, 1976), должны действовать тормозящим образом на систему теплообразования и вызывать снижение темперачуры тела. Степень торможения и снижения температуры в первую очередь зависит от исходного уровня: чем выше температура от нормального предела, тем более выраженным будет эффект рефлекторного воздействия. Это общий принцип. Рефлекторный эффект действует только когда есть сдвиг, когда возбудимость либо выше, либо ниже исходного. Поэтому при нормальной температуре тела воздействие на точки акупунк-туры не дает эффекта снижения. Напротив, при порышен-ной температуре хорошо выражен рефлекторный эффект снижения. Методика работы должна быть такой же, как при обезболивании или лечении болевых синдромов. Целесообразность снижения температуры при некоторых заболеваниях мы разбирать не будем, так как этот вопрос решает врач каждый раз отдельно, с учетом многих других условий. Нам важно подчеркнуть принципиальную возможность рефлекторного воздействия на терморегулирующую систему организма через мезенцефалические и диэнцефаличе-ские механизмы.
Чтобы завершить разбор физиологических механизмов участия гипоталамуса в регуляции внутренней среды организма, стабильность которого является непременным условием для его осуществления, как подчеркивал еще Клод Бернар (1937), необходимо обратиться к гипоталамо-гипофизарным взаимоотношениям и к механизмам нейро-эндокринных регуляций.
Рефлексотерапия, в частности аурикулярная рефлексо-терапия, является мощным инструментом воздействия на эти механизмы с целью их нормализации. Между гипо-таламчческими ядрами и гипофизом существует тесная связь .(рис. 23). Гипофиз — это, по-существу, важнейшая часть гипоталамуса, его химический комбинат, которому он поставляет полуфабрикаты (предшественники гормонов) и программу действий (нейро-гуморальные рогуля-горные эффекты), в зависимости от потребностей организма. Получая информацию о состоянии внутренних органов и внутренней среды организма от окружающих интегративных центров мозга (ретикулярной формации, лимбической системы, таламо-кортикальной системы), гипоталамус формирует адекватные для ситуации эффекты управления, которые реализуются: а) нейрогенным путем, через гипотала-мические связи со стволом мозга на эффекторные центры симпатических и парасимпатических систем и далее по соответствующим ретикуло-спинальным путям и нервам на органы-исполнители; б) нейро-гуморальным путем на гипофиз и его гормональную функцию, который в свою очередь действует на другие эндокринные железы и через их гормональную функцию на органы и системы организма.
Гипоталамо-гипофизарная система в первую очередь ответственна за три основные группы гормонов: ГТГ— гонадотропные, АКТГ — адренокортикотропные и ТТГ— тиреотропные (Я. Сентаготаи, Б. Флерко, Б. Меш, Б. Ха-лас, 1965; К. Лишшак, Э. Эндрёци, 1967). выделяемые передней долей гипофиза.
В гипофизе четко выделяются две большие дольки:
передняя — аденогипофиз и задняя — нейрогнпофиз. Имеется также средняя или промежуточная долька — значительно меньшая по размерам, которую чаще относят к задней доле гипофиза. Роль промежуточной дольки долгое время оставалась невыясненной. Известно, что там образуется гормон, стимулирующий образование меланоцитов,интермедии который регулирует изменение окраски кожи, воздействуя на меланофоры. У человека интермедии имеет значение в регуляции клеток пигментного слоя в глазу. При ярком свете клетки пигментного слоя выпускают псевдоподии, благодаря чему задерживают избыточное проникновение световых лучей к сетчатке глаза и предохраняют ее от чрезмерного раздражения.
После открытия эндогенных опиатов промежуточная долька гипофиза рассматривается как депо для предшественников эндорфинов (см. ниже).
Задняя доля гипофиза, получившая название — нейро-гипофиза, связана с гипоталамусом при помощи гипотала-мо-гипофизарного нервного тракта, образованного аксонами крупных нейронов супраоптического и паравентрику-лярного ядер гипоталамуса, напоминающих секреторные клетки (рис. 23). Благодаря аксоплазматическому току осуще
|
из
5.00
|
Обсуждение в статье: Ретикулярная формация и гиго галамус. Висцерально-эндокринные функции |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы