 |
Главная |


Функциональные особенности нервов и нервных центров ушной раковины
|
из
5.00
|
Теперь, когда мы познакомились с эволюционными основами нервных приборов ушной раковины, следует более подробно рассмотреть функциональные характери-стихи отдельных нервов уха и особенности их центральной организации.
Какими бы богатыми и разное Зразными не были нервы ушной раковины и как бы важна не была их роль в эволюции, главным для понимания потенциальных возможностей аурикулярных рефлексов у человека на различные функции организма являются внутренние связи аурикулярных нервов, центры, которые они активируют и включают в осуществление тех или иных рефлексов.

Рис. 12. Схема иннервации лица, головы и шеи.
Условные обозначения:
V - верхнечелюстная (максиллярная) ветвь V нерва; Vg—глазная (оф-тальмическая) ветвь V нерва; Vg — нижнечелюстная (мандибулярная) ветвь V нерва, образующая ушные нервы; С, — С4 — нервы шейного сплетения.
Важно показать, что богатое наследство, накопленное в процессе эволюции, в основном сохранилось, хотя и с заметными изменениями, и может быть использовано у человека достаточно эффективно.
Тройничный нерв—п. trigemi-nus (V пара) является самым большим краниальным нервом и хотя принадлежит к смешанным нервам, однако является преимущественно сенсорным и содержит мощный пучок афферентных волокон от различных участков лица и передней половины головы (рис. 12). Моторная ветвь иннервирует жевательные мышцы и небольшое моторное ядро, расположенное в стволовой части мозга, в области варо-лиева моста, рядом с главным сенсорным ядром тройничного нерва (рис. 13). Тройничный нерв имеет три крупных сенсорных ядра и большой периферический сенсорный ганглий (полулунный или гассеров узел). Число ядер тройничного нерва, хотя и совпадает с числом трех основных ветвей этого нерва, однако это лишь случайное совпадение, так как три ядра трои-' ничного нерва не предназначены каждое для трех различных его ветвей, а каждое из них имеет свое специальное назначение и связаны они по-разному с ветвями тройничного нерва.
Главное сенсорное ядро (рис. 13) располагается в тег- j ментальной области варолиева моста и является, пс-суще-

Рис. 13. Схема распределения сенсорных ядер афферентов ушной раковины в стволе мозга.
Справа — на горизонтальном плане.
Слева— на фронтальных планах, соответствующих срезам А—Г, указанным стрелками на горизонтальном плане.
Условные обозначения: I — мезенцефалическое (восходящее) ядро V нерва; 2 — главное сензорное ядро V нерва; 3 — спинальное (нисходящее) ядро V нерва; 4— солитарное ядро V, VII, IX, Х нервов;
5—обоюдное ядро (п. ambiguus); 6 — двигательное ядро V нерва;
7 — двигательное ядро VII нерва; 8 — центральное серое вещество;
9 — нежный (gracilis) канатик и нежное ядро дорзального столба спинного мозга; 10— клиновидный (cuneatus) канатик; 11 — клиновидное ядро дорзального столба спинного мозга; 12 — нисходящий (спиналь-ный) тракт V нерва; СМ — средний мозг; ВМ— варолиёв мост, ПМ — продолговатый мозг (границы обозначены двумя черточками); А, Б, В, Г — места фронтальных срезов.
ству, гомологом нежного и клиновидного ядер дорзальных канатиков спинного мозга. Хотя сенсорная система тройничного нерва в общих чертах организована так же, как спинальная система, однако имеются и некоторые особенности, которые придают тройничной системе исключительное значение в афферентных функциях центральной нервной системы. Главное сенсорное ядро получает информацию от покровов лица, головы, полости носа, рта, от ушной раковины и передает эти сигналы непосредственно в таламус при помощи мощного пучка быстропроводящих волокон, которые идут вместе с волокнами медиального лемниска и заканчиваются в специфическом сенсорном реле таламу-са — заднем вентральном ядре (VP).
После серии работ Mountcastle (1961) и его сотрудников (Rose, Mountcastle, 1959), а также специальных исследований, выполненных нами (Р. А. Дуринян, 1965) и Ю. П. Лиманским (1976) стало очевидным, что медиальный полюс заднего вентрального ядра таламуса, то есть вентральное задне-медиальное ядро (VPM) является, по-существу, представительством чувствительности лица и головы; здесь частично представлена также чувствительность передней (верхней) конечности (рис. 28). На это обстоятельство надо обратить внимание, так как оно свидетельствует об исключительной роли афферентов области лица и головы (тройничный нерв, шейное сплетение, нервы ушной раковины) в активации структур таламуса и коры больших полушарий, не говоря уже о первичной активации стволовых структур.
Из главного сенсорного ядра тройничного нерва в области варолиева моста (рис.11) в таламус, в ядро VPM направляются как перекрещенные, так и прямые (неперекрещенные) проекционные афферентные волокна. Это обстоятельство указывает на то, что при воздействии на различные области головы, лица, ушной раковины, иннервируемые афферентами тройничного нерва, сенсорные сигналы, одновременно направляясь в оба таламуса, активируют синхронно и правое и левое VPM. В свою очередь по соответствующим таламо-кортикальным проекциям синхронно. активируются сомато-сенсорные области коры в обоих полушариях мозга и ряд подкорковых структур (рис. 14). Следовательно, стимуляция точек акупунктуры в области лица и головы, как и стимуляция точек акупунктуры ушной раковины, на одной стороне может вызывать синхронную активацию сенсорных систем обоих, не только контралате-рального, но и ипсилатерального, полушарий и включать врефлекторную реакцию билатеральные механизмы мозга (см. ниже).
Основной поток искусственно вызванных сенсорных сигналов при стимуляции точек акупунктуры, принадлежащих тригеминальной системе (в том числе на ушной раковине), направляется после перекрестка в противоположный таламус, как это и должно быть в отношении любых афферентных проекций. Однако эти сигналы поступают не только в таламическое ядро VPM, но и в другие ядра, а

Рис. 14. Схема окончаний тригеминальных афферентов в вентральном заднемедиальном ядре таламуса и их проекции в коре.
Слева — перекрещенные; справа — неперекрещенные
MD — медиодорзальное ядро
VPL — вентральное задне-латеральное ядро
VPM — вентральное задне-медиальное ядро
затем и в кору несколько позже (на несколько миллисекунд), чем сигналы, идущие по короткому и быстрому пути из главного тригеминального сенсорного ядра. Это объясняется тем, что второй путь, который во многом напоминает систему спино-таламического тракта, начинается от другого ядра тройничного нерва.
Нисходящее или спинальное ядро тройничного нерва (см. рис. 10, 11 и 13)—самое протяженное сенсорное ядро ствола мозга и располагается своим верхним концом в тегментальной части варолиева моста, а нижним достигает четвертого шейного сегмента спинного мозга. Это ядро является гомологом чувствительных ядер задних рогов спинного мозга, что указывает на функциональную общность тригеминальных афферентов со спинальными и сви-детельствуют о том, что такие анатомические понятия, как краниальный нерв, носят скорее условный характер. Тройничный нерв, как и другие краниальные нервы, иннервиру-ющие покровы тела, то есть афферентные сомато-висцераль-ные нервы общей чувствительности можно рассматривать как более специализированные спинальные нервы, которые сильно исказили или даже утратили свою сегментарность на периферии, но сохранили общий принцип центральной организации. Другое дело, краниальные нервы специализированных органов чувств, которые действительно утратили свою сегментарность полностью, которые имеют свои самостоятельные ядра в таламусе и самостоятельные корковые проекции (зрение, слух и др.).
Тройничный нерв, нервы шейного сплетения, как и другие нервы ушной раковины, следует рассматривать под двумя ракурсами. С одной стороны,— выделяя то общее, что их объединяет как афферентную систему общей чувствительности ушной раковины, и с другой,— выделяя то, что является специфичным для каждого из афферентов.
Общим для всех афферентных нервов ушной раковины является то, что передаваемая ими информация направляется в одно и то же ядро таламуса — заднее вентро-медиаль-ное ядро (VPM). Здесь нет принципиальных различий в принадлежности к спинальной или краниальной системе;
проекции всех афферентных нервов в ядре VPM организованы одинаково (Р. А. Дуринян, 1965). Мы рассмотрим этот вопрос специально, а сейчас продолжим анализ функциональных особенностей нервов ушной раковины.
В спинальном ядре тройничного нерва заканчиваются преимущественно наиболее тонкие афферентные волокна температурной и ноцицептивной (бэлевой) чувствительности. От чувствительных клеток этого ядра начинается проекционный путь в ядра таламуса, однако в отличие от проекций волокон вторых нейронов главного чувствительного ядра тройничного нерва, где заканчиваются наибэлее толстые афференты тактильной чувствительности, аксоны вторых нейронов спинального ядра дают проекции не только в ядро VPM, но и в группу неспецифических ядер таламуса (срединный центр — СМ и другие ядра интраламинарной группы).
Сигналы, передаваемые этими волокнами, активируют не только локальные специфические проекции вентрального таламуса, но и диффузные, мультисенсорные нейроны с так называемыми неспецифическими проекциями в коре иподкорковых ядрах. Роль неспецифических систем вмеханизмах мозга мы рассмотрим отдельно.
Следовательно, чтсбы активировать неспецифическую систему таламуса, что равносильно включению в рефлекторный процесс множества структур мозга, необходимо наносить на точки акупунктуры тригеминальной системы (и других областей тела), в том числе ушной раковины, воздействия достаточной интенсивности, которые будут восприниматься на пороге болевых. Конечно, они должны быть легко переносимыми для больного, чтобы не быть пыткой для него, но тем не менее достаточно интенсивными для активации высокопороговых «С» и А-дельта.
Весьма эффективными для активации высокопороговых волокон, помимо интенсивных механических воздействий иглами ноцицептивного порога, должны быть локальные термические раздражения ушной раковины (и других областей тригемкнальной системы), которые в практике ау-рикулярной рефлексотерапии используются крайне мало.
Для термостимуляции точек ушной раковины можно применять сфокусированный луч какого-либо электрического источника тепла (лампочка, спираль накаливания, полупроводниковый элемент и т. п.), интенсивность теплопродукции которого можно легко регулировать. Однако возбуждение высокопороговых терминалей тройничной системы, как и любой другой, на ушной раковине (ноцицеп-тивных и термических) может быть достигнуто весьма эффективно не только натуральными способами раздражения (механическим — болевым или тепловым), но и при помощи электрических импульсов. Электрические стимулы по сравнению со всеми другими известными в настоящее время способами физического воздействия на возбудимые ткани организма обладают огромным преимуществом, которое заключается в том, что электрические потенциалы легко управляемы по амплитуде, форме, частоте, длительности. Электрический ток является адекватным раздражителем для возбудимых тканей и не дает последействия;
в разумных пределах совершенно безопасен и может применяться любому больному, в любом возрасте. Электрические стимулы можно наносить через кожу, без необходимости дополнительного введения игл в точки акупунктуры или в места скопления нервных элементов, что не только упрощает процедуру, но делает ее еще более безопасной.
Не случайно в мировой практике электростимуляция точек акупунктуры завоевывает все более широкое призна-ние и постепенно вытесняет классический метод игл, даже на родине метода.
Итак, активация высокопороговых волокон тригеми-нальной системы в ушной раковине, так и в других областях, может быть достигнута вполне адекватной электростимуляцией, которая в ближайшие годы, по-видимому, полностью вытеснит из практики классические иглы, как массовое явление в рефлексотерапии.
Все, что было сказано в отношении принципов активации терминален тройничного нерва, имеет абсолютное отношение и к другим нервным проводникам , так как основано на функциональных законах электровозбудимости нервных волокон, начало которых положили работы Э. Дюбуа-Реймона, К. Пфлюгера, Н. Е. Введенского, X. Гассера и многих других (Brazier, .1968).
Афферентные волокна тройничного нерва, заходя в варолиев мост на уровне главного сенсорного ядра, разветвляются на два основных тракта. Первый, образованный из толстых, быстропроводящих и низкопороговых волокон, как уже отмечалось, оканчивается в этом ядре (рис. 11). Второй, образованный главным образом из тонких, медлен-нопроводящих и высокопороговых волокон типа А-дельта и С образует нисходящий спинальный тракт нерва, который заканчивается на нейронах одноименного ядра. Интересно, что волокна третьей — мандибулярной ветви тройничного нерва, откуда образуется также аурикулярная ветвь тройничного нерва (п. auriculo-temporalis), опускаясь вниз, достигают четвертого (иногда даже пятого) шейного спи-нального сегмента. Вместе с нисходящими волокнами мандибулярного нерва располагаются также (см. рис. 10 и 11) нисходящие волокна VII, IX, Х пар черепно-мозговых нервов (Crosby, Hamphrey, Lauer, 1962; Curtis, Jacobson Marcus, 1972), которые заканчиваются в том же тригеми-нальном ядре, и которые далее образуют такие же проекции в неспецифических ядрах таламуса, как и высокопороговые тригеминальные волокна. Наконец, как уже упоминалось, на нейронах спинального, тригеминального ядра контактируют афферентные волокна спинальных сегментов, в частности тех (Cg—Сз), волокнами которых образованы шейные нервы ушной раковины (п. auricuraris magnus, п. occipitalis minor).
Следовательно, в отношении информации, передаваемой высокопороговыми волокнами термической и ноцицептив-ной чувствительности в нисходящее спинальное ядро нетпринципиальных различий для нервов шейного сплетения, для нервов VII, IX, Х пары или для тригеминальных веточек ушной раковины. Разница может быть лишь в пространственной локализации, но это уже функция толстых низкопороговых волокон, которые имеют свои первичные ядра в стволе мозга. Для быстропроводящих волокон шейных нервов это клиновидное ядро продолговатого мозга; для тройничного нерва—главное сенсорное ядро в варолиевом мосту; для нервов VII, IX, Х пары — солитар-ное ядро в продолговатом мозгу и варолиевом мосту (рис. 10, 11 и 13). Вторичные волокна от этих ядер идут в восходящем направлении, совершают перекрест и в составе медиального лемниска направляются в таламус.
Третьим сенсорным ядром тройничной системы является мезенцефалическое ядро, которое расположено лате-рально от четвертого желудочка и сильвиевого водопровода на уровне верхней половины варолиева моста и среднего мозга. Хотя это ядро меньше, чем спинальное ядро, но все же заметно больше клиновидного и нежного ядер дорзальных канатиков спинного мозга. Такое ядро получает преимущественно проприоцептивную информацию по наиболее толстым низкопороговым волокнам от мимических и жевательных мышц. Здесь важно вспомнить, что манди-булярная ветвь тройничного нерва образует ушно-височный нервы, который иннервирует ушную раковину и височную область, в частности, височную мышцу и сустав нижней челюсти. В связи с этим не исключена возможность, что ушная ветвь данного нерва может содержать волокна, которые будут активировать мезенцефалическое ядро тройничного нерва. Большие размеры мезенцефалического ядра тройничного нерва по сравнению с клиновидным ядром задних канатиков указывают на то, что количество афферентных нервных волокон, контактирующих на нейронах этого ядра, значительно больше, чем количество волокон, идущих в клиновидное ядро от шейного (рис. 13) и частично плечевого сплетений, в то время как кожная поверхность, иннервируемая тройничным нервом, заметно меньше, чем поверхность кожи, иннервируемая шейным (Ci—C.,) и плечевым сплетением (Сз—ТП^), которое формируется нервами шейных сегментов спинного мозга.
Афферентная система тройничного нерва является самой мощной из всех соматических афферентов Организма и обладает огромными потенциальными возможностями для активации специфических и неспецифических структурствола мозга и таламуса и соответствующих корковых проекций. Тесную морфо-функциональную связь с системой тройничного нерва имеет шейное сплетение (Ci—С.,) и чагтлчнэ плечевое сплетение (Сд—Thi). Это отчасти объясняется некоторыми особенностями механизмов нервной рггуляцчи движений головы и особенно эмоциональной мимикой лица и эмоциональными позами, связанными с дв1ж;ниям'-1 головы и верхних конечностей. Для осуществления рефлекторных движений с целью поддержания равнэзгсля прл отклонении головы и для всего комплекса позных и мимических двигательных реакций в ответ на какэе-либэ эмоциональное состояние (страх, радость, гнев, удэвэльствче и т. п.) необходима четко скоординированная и согласованная работа различных мышечных групп об-ла:тл л та (м-шлка), шеи (постановка головы), рук (жесты). Для этого тргбуется богатая и тонко дифференцированная афферентная информация от поверхностных (рецепторы кожи) и глубоких (проприоцепторы) структур указанных областей, которая интегрируется в первичных стволовых сенсорных ядрах этих афферентных систем и в стволовой ретикулярной формации для обеспечения автоматических реакций. Для более сложной координации движений и быстрой адаптации их к постоянно меняющимся условиям внешней среды, то есть для правильной оценки ситуации и формирования адекватной приспособительной реакции, эти сигналы переключаются со стволового уровня (уровня автоматических реакций) на таламо-корковый уровень {уровень адаптивных реакций).
Мы уже указывали на существование коллатеральных связей между афферентными волокнами тригеминального и шейного нервов в спинальном ядре V нерва (рис. 11);
вполне возможны такие связи и на уровне главного и ме-зенцефалического ядер тройничного нерва с волокнами шейных нервов. Это создает благоприятные возможности для взаимодействия сигналов, поступающих в указанные ядра по шейным и тригеминальным афферентам. Из нервов плечевого сплетения наибольшую близость с тригеминаль-ной системой имеет пятый шейный корешок (Q), так как известны коллатерали афферентных волокон этого корешка, связанные с нейронами спинального ядра V нерва (Crosby, Hamphrey, Lauer, 1962). Это важно, так как Cs участвует в формировании лучевого нерва (п. radialis), чувствительные волокна которого в составе поверхностного лучевого нерва (п. radialis superficialis) иннервируют кожу в обла-сти большого, указательного и среднего пальцев руки (рис. 15). Лучевой нерв является самым большим из нервов плечевого сплетения; его афферентная часть очень развита, так как обеспечивает чувствительную иннервацию наиболее важных пальцев руки; тех пальцев, которые у

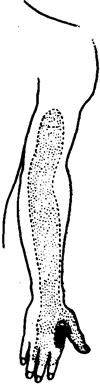
Рис. 15. Схематическая топография лучевого нерва (слева) и область чувствительной иннервации кожи (очерчена пунктиром и отмечена точками^ веточками поверхностного лучевого нерва.
Слева—стрелкой указана область разветвления поверхностного лучевого нерва, которая совпадает с акупунктурной точкой 4 GI (хэ-гу);
Справа— частыми точками указана область наиболее интенсивной чувствительной иннервации кожи, которая совпадает с проекциями точки 4 GI (хэ-гу)
человека в функциональном отношении наиболее дифференцированы, которые выполняют самые тончайшие движения, на какие только способна рука человека. В связи с этим информация, передаваемая по лучевому нерву, активирует множество систем мозга (например, зрительный контроль движений пальцев) и требует участия самого большого (как и рече-двигательная функция) количества сенсомо-торных нейронов коры больших полушарий. Вот почему гомункулюс—и сенсорный и моторный—в коре имеет огромные (по сравнению с другими частями тела) пальцы, и огромное лицо (рис. 3). В этом отношении лучевой нерв — его афферентная часть весьма сходна с мандибулярной ветвью тройничного нерва, который своими проприоцеп-торами участвует в афферентном контроле жевательной и рече-двигательной функций.
Функциональная дифференцированность пальцевого и -рече-двигательного аппарата и функциональная общность
центральной организации этих афферентных систем очевидны, хотя бы из такого важного факта, что афферентные проекции тройничного нерва представлены (рис. 28) на таламическом уровне в одном и том же заднем вентро-ме-диальном ядре (VPM), в то время как чувствительность всего остального тела занимает заднее вентро-латеральное ядро (VPL). На таламическом уровне, в VPM благодаря широкому перекрытию между афферентными проекциями области лица, головы, шеи и верхних конечностей имеются возможности для интенсивности взаимодействия сигналов, конвергирующих на одни и те же нейроны. Физиологические возможности такого взаимодействия будут рассмотрены специально, а сейчас необходимо подчеркнуть, что на стволовом, таламическом и корковом уровнях афферентные сигналы, вызванные в волокнах лучевого нерва, могут активировать не только собственные проекции, но и элементы тригеминальной системы.
На рисунке 15 видно, что место разветвления поверхностного лучевого нерва на нервы пальцев точно совпадает с одной из наиболее популярных и эффективных точек акупунктуры — с точкой хе-гу (четверта я точка, так называемого меридиана толстой кишки — 4GI).
Небольшой участочек кожи над этой точкой (рис. 15) особенно интенсивно иннервирован афферентами лучевого нерва (Chusid, 1970). Интересно, что этот участочек располагается как раз в области проекции точки хе-гу. Не исключена возможность, что рефлекторная «мощь» точки хе-гу объясняется концентрацией афферентных терминалей лучевого нерва в покровах межпальцевой ямки, но главное, конечно, в особенностях центральной организации афферентных проекций области кисти и потенциальной возможности вовлечения в активность обширной системы стволовых и таламокортикальных проекций.
Принадлежность точки хе-гу к лучевому нерву признают Bossy (1975), Bossy, Godlewsgu, Maurel (1975), Kaada <1976) и другие авторы.
На примере центральной организации проекций лучевого нерва и этой точки мы хотели продемонстрировать, что при стимуляции какой-либо точки, в том числе на ушной раковине, рефлекторная реакция не должна рассматриваться как локальная, ограниченная в пределах проекции нескольких афферентных волокон, что в зависимости от функциональных особенностей той или иной афферентной системы и ее внутрицентральных связей рефлекторные ре-акции могут носить более многообразный и сложный характер, взаимодействуя с различными системами.
При воздействии на точки ушной раковины в области тригеминальной иннервации активация развивается не только в тригеминальных ядрах и их последующих проекциях, но может охватывать системы проекций шейного и плечевого сплетения, а также ряда проекций висцеральных афферентов.
Вместе с нисходящими волокнами мандибулярной ветви (Уз) тройничного нерва в продолговатом мозге располагаются также нисходящие волокна VII, IX и Х пар краниальных нервов. Все эти нисходящие аксоны, как и тригеминальные, прерываются на нейронах спиналыюго ядра. V нерва (рис. 10), после чего подавляющее большинство вторичных аксонов совершает перекрест, принимает восходящее направление и наподобие спино-таламическому тракту присоединяется к медиальному лемниску и заканчивается в неспецифических ядрах таламуса. Следовательно, функциональная организация центральных проекций афферентных систем, образованных высокопороговыми волокнами нервов VII, IX и Х аналогична проекциям таких же волокон V нерва. Различие может касаться лишь количественной стороны вопроса в связи с тем, что тригеминаль-ный нерв является самым большим афферентным коллектором, то есть состоит из значительно большего количества волокон. Однако аналогично не означает равносильно, так как каждая афферентная система имеет и свои характерные особенности, главным из которых является то, что афферентный сигнал прежде всего включает в реакцию систему прямых рефлекторных связей.
Лицевой нерв (VII пара) в отличие от тройничного является преимущественно моторным. Сенсорная или афферентная ветвь его значительно меньше и образована промежуточным нервом Врисберга; разделяется на два небольших нерва: ушной и язычный. В эволюции этот нерв претерпел большие изменения, сильно утратив свою афферентную часть за счет тройничного нерва и, по-видимому, шейных нервов. То, что осталось от афферентной системы лицевого нерва у человека в виде промежуточного нерва, это ветви, которые иннервировали хеморецепторы ротовой полости и жаберных мешков у рыб.
Одна ветвь так и сохранила свою хеморецептивную принадлежность в виде язычного нерва, который иннервирует вкусовые сосочки передней части языка.
Вторая ветвь, которая образует ушную ветвь, утратила хеморецептивную специфичность и трансформировалась it систему общей чувствительности.
Афферентные волокна лицевого нерва, после периферического ганглия, поступают в латеральную часть ретикулярной формации на уровне варолиева моста и направляются в ядро солитарного тракта, которое располагается непосредственно рядом с центральной (периакведуктальной) серой массой, у дна четвертого желудочка. Рядом с этим ядром несколько латерально расположены все три ядра тройничного нерва, особенно полно мезенцефалическое ядро (рис. 10). Солитарное ядро является одним из важнейших сенсорных узлов в стволовой части мозга. Помимо афферентов лицевого нерва, которые входят в ростральный отдел ядра, здесь же заканчиваются также волокна IX и Х нервов, причем языко-глоточные афференты располагаются каудально, после афферентов лицевого нерва, а всю остальную и большую часть ядра занимают афференты блуждающего нерва. В солитарном ядре заканчиваются и некоторые афференты тройничного нерва (Е. К. Сепп, 1959), хотя, какие именно, точно не установлено.
Язычная ветвь имеется, как известно, и в составе языко-глоточного нерва, что указывает на явную функциональную общность хеморецепторных афферентов VII и IX нервов, которые сформировались в эволюции на стадии рыб.
У человека мы можем отметить явные элементы общности в функциональных, эволюционных и анатомических особенностях организации афферентов лицевого и языко-глоточного нервов, связанных с вкусовой рецепцией. С другой стороны, оба нерва, как и блуждающий, имеют ушные ветви и иннервируют область конхи, трехсторонней выемки и ряд близлежащих областей уха. Учитывая небольшую относительную мощь этих афферентов по сравнению с тройничными и шейными афферентами на ушной раковине и, с другой стороны, наличие хеморецептивных (вкусовых) афферентов в составе VII и IX язычных нервов,. которые заканчиваются, как и ушные афференты VII, IX и Х нервов в солитарном ядре, мы предложили специальный физиологический прием усиления рефлекторного эффекта стимуляции области конхи, особенно полости конхи (cavum conchae), где нет соматических афферентов, в отличие от трехсторонней выемки.
Прием этот заключается в том, что перед тем как воздействовать на область конхи (иголками, электрическимистимулами, фокусированным световым лучом и т. п.) больному дается на язык кисло-сладкая таблетка или драже (лучше всего аскорутин; можно кусочек сахара, сахарина, леденец и т. п.). Благодаря воздействию на вкусовые рецепторы кончика языка (VII нерв), а затем корня языка (IX нерв) возникает распространенная волна возбуждения в хеморецептивных афферентах языка, которая достигает солитарного ядра и вызывает там диффузную активацию определенной группы нейронов. На эти нейроны с повышенной возбудимостью затем поступают сигналы, вызванные с локальных пунктов ушной раковины, в результате чего их ответная реакция оказывается усиленной. Некоторые испытуемые отмечали, что на фоне вкусового раздражения слабые электрические стимулы, наносимые на область конхи, воспринимаются субъективно, как более интенсивные, чем эти же стимулы без предварительного раздражения вкусовых рецепторов. Следовательно, искусственно создавая повышенный фон возбудимости нейронов солитарного ядра, можно усилить результат аурикулярных рефлексов. Нечто подобное имеет место при корпоральной акупункту-ре при стимуляции общих тонизирующих точек.
Nogier (1972) предлагает стимулировать нулевую точку ушной раковины в тех случаях, когда рефлекторная реакция слабо выражена как при стимуляции точек уха, так и при определении отраженных болей на ушной раковине. Стимуляция нулевой точки уха для усиления рефлекторного эффекта, по-видимому, играет такую же роль предварительного изменения возбудимости нейронов релейных структур ствола мозга (тригеминальные ядра, солитарное ядро, чувствительные ядра задних рогов шейных спиналь-ных сегментов и другие), как и стимуляция тонизирующих точек тела.
Какое значение имеет фон возбудимости нейронов центральной нервной системы для характера их ответных реакций, мы рассмотрим при обсуждении нейрональных механизмов рефлекторных реакций. А пока продолжим рассмотрение центральной организации афферентных проекций аурикулярных нервов.
Вторичные аксоны солитарного ядра идут в восходящем направлении (наподобие тригеминальных), входят в состав медиального лемниска и заканчиваются в том же заднем вентро-медиальном ядре таламуса. Следовательно, сенсорная информация от солитарного ядра поступает в то же таламическое реле, куда приходят сигналы от сенсорныхядер тройничного нерва, а также от шейного сплетения через клиновидное ядро продолговатого мозга (рис. 28).
При стимуляции ушной раковины, например, у начала корня завитка, где представлены афферентные терминали V, VII, IX и Х нервов (см. рис. 7,6), волна возбуждения распространиться в солитарное ядро и в тригеминальные ядра. Кроме того, нисходящие спинальные волокна манди-булярной ветви тригеминального нерва и некоторые волокна VII, IX, Х нервов, идущие в составе этих нисходящих волокон (рис. 10), достигая четвертого спиналь-ного сегмента, вовлекают в активность верхний шейный отдел спинного мозга. Таким образом, при локальной стимуляции определенных точек ушной раковины волна возбуждения появляется сразу же в нескольких ядрах ствола мозга, и каждое ядро посылает восходящий активирующий поток сигналов в одно и то же специфическое ядро таламу-са (VPM). Однако дело не исчерпывается этим. Одновременно поток сигналов из ядер ствола направляется в неспецифические ядра таламуса. Наконец, параллельно с активацией таламических структур происходит активация многих структур ретикулярной формации ствола мозга, откуда волна возбуждения передается в восходящем направлении в неспецифический таламус, в ядра гипоталамуса, в лимбическую систему, и, конечно, в кору больших полушарий.
С другой стороны, при возбуждении ретикулярной формации ствола могут возникать непосредственные нисходящие эффекты (ретикуло-спинальная система) на спиналь-ный сегментарный аппарат, что может повлиять на моторные и сенсорные реакции спинальных структур мозга. Ретикулярная формация ствола мозга со своей стороны находится под контролем вышележащих структур, особенно коры больших полушарий. Благодаря этому волна возбуждения, вызванная стимуляцией ушной раковины при поступлении в корковые проекции, может включить механизм коркового контроля, который изменит весь характер реакции. Вот почему совершенно прав Nogier (1972), когда рекомендует приступать к сеансу аурикулотерапии только после того, как больной успокоится, то есть снизится избыточное возуждение мозга, когда механизм коркового контроля не будет экзальтирован, и, следовательно, не будет извращать сложные, многоступенчатые рефлекторные реакции. Эти вопросы, как и нейрональные механизмывзаимодействия различных сигналов, требуют отдельного анализа.
Данный раздел завершим рассмотрением последнего из аурикулярных нервов — блуждающего нерва (X пара краниальных нервов)—п. vagus. Как следует из названия,. этот нерв является самым протяженным и распространенным нервным каналом из всех краниальных нервов. Сенсорным ядром Х нерва, как уже известно, является солитарное ядро, которое располагается рядом с нисходящим тригеминальным ядром. Между ними расположены два моторных или эффекторных ядра Х нерва: дорзальное ядро и обоюдное ядро — п. ambiguus (рис. 13). Между солитарным ядром и эффекторными ядрами блуждающего нерва имеются прямые двухнейронные связи, что обеспечивает возможность быстрых рефлекторных реакций в пределах органов, иннервируемых эффекторами блуждающих нервов, а к таковым относятся почти все внутренние органы, за исключением тазовых (рис. 16). В связи с тем, что солитарное ядро является первичным сенсорным реле не только

| Рис. 16. Внутренние органы, иннервируемые блуждающим нервом. Нижняя треть толстого кишечника и тазовые органы иннервнруются тазовым нервом. |
для афферентов вагуса, но и VII и IX нервов, а также некоторой части тригеминальных волокон (Е. К. Сепп, 1959), непосредственные рефлекторные реакции могут быть вызваны во внутренних органах и системах организма, контролируемых вагусом, при возбуждении афферентов этих нервов, а они, как известно, иннервируют ушную раковину.
Заключение
Прямые рефлексы по типу: «стимул-реакция» не могут Сыть достаточно продолжительными и, следовательно, эффективными для лечения серьезных патологических отклонений. Рефлексы подобного типа при однократном применении могут успешно использоваться лишь для воздействия на быстротекущие процессы. Например, для кратковременного купирования спастического состояния гладкой мускулатуры или для усиления секреции, мышечных сокращений. При повторных многократных воздействиях, если причина спазма имеет центральное (нейрогенное) происхождение, можно полностью купировать это состояние благодаря тому, что блокируется очаг пароксизмаль-ного возбуждения в определенной группе центральных нейронов, которые управляют тонусом гладкой мускулатуры данного органа или области.
При более серьезных формах патологии механизм терапевтического эффекта не исчерпывается простым рефлексом «стимул-реакция» на уровне двухнейронной рефлекторной дуги, а реализуется благодаря участию целого ряда других структур мозга, в том числе вышележащих структур, которые управляют (контролируют) эфферентным выходом на данную функцию или систему.
Все внутренние органы, вовлеченные в систему вагусной иннервации, снабжаются одновременно и афферентными и эфферентными волокнами, что и обеспечивает надежную рефлекторную связь внутри этой системы. Рефлекторные влияния на функции внутренних органов и систем могут запускаться через другие афферентные входы, которые тесно взаимодействуют с системой афферентов вагуса непосредственно на уровне нейронов первичного реле — соли-тарного ядра, как например, афферентыУП, IX и Х нервов. Такое взаимодействие более сложного характера может иметь место в ретикулярной формации и на таламическом уровне, так как вторичные аксоны афферентной системы вагуса после переключения в солитарном ядре принимают восходящее направление, перекрещиваются, присоединяются к медиальному лемниску и заканчиваются, как и три-геминальные афференты, в таламическом ядре VPM (рис 28). Эта проблема была подробно разработана нами ранее и представлена в отдельной публикации (Р. А. Дуринян, 1965).
Таким образом, в VPM заканчиваются вторичные аксоныафферентов V, VII, IX и Х нервов, а также шейного сплетения и все они участвуют в иннервации уха. Следовательно, чувствительность ушной раковины представлена в таламическом яд
|
из
5.00
|
Обсуждение в статье: Функциональные особенности нервов и нервных центров ушной раковины |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы