 |
Главная |


Принцип организации фагмид (например, pBluescript).
|
из
5.00
|
Фагмиды - это плазмидные векторы, образованы внедрением в векторы pUC фрагмента ДНК фага f1 или М13, содержащие ori+ и ori- нитевидных фагов, а также pac-сайт (необходим для морфогенеза фаговых частиц). Рекомбинантные фагмиды распознают по изменению окраски негативных колоний на чашках с X-gal.
Строение векторов Bluescript M13- и Bluescript M13+:
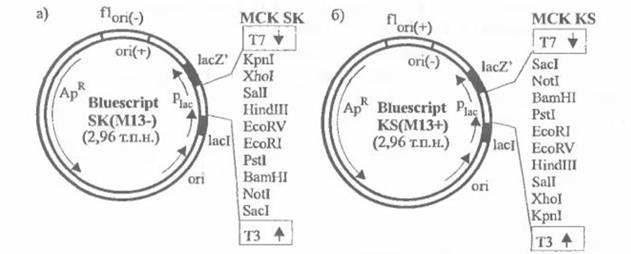
Векторы M13- и M13+ отличаются направлением ori-сайта, взятого у фага f1, и тем, какая нить вектора будет упакована в фаговую корпускулу.
Буквы SK (SacI→KpnI) и KS (KpnI→SacI) в названии вектора указывают на ориентацию сайта поликлонирования MCS. Стрелки в MCS указывают на направление транскрипции с фаговых промоторов Т3 и Т7.
В присутствии фага-помощника f1(или M13) фагмиды, используется сайт ori+, начинают реплицироваться как фаговые ДНК, образуя однонитевые копии плазмид, причем выбор нити для копирования зависит от ориентаии ori-сайта. Образовавшиеся однонитевые ДНК могут упаковываться in vivo в капсулу и одновременно с фагом-помощником покидать клетку естественным путем. В препаратах однонитевые ДНК, выделенных из фагов, присутствует смесь ДНК фага-помощника и фагмид. Доля фагмидной ДНК в препаратах зависит от структуры ori-сайтов фагмиды и фага-помощника.
Таким образом, с помощью фагмид клонируемый фрагмент может быть получен и использован в одно- или двунитевой форме. Их преимущество перед векторами M13mp – возможность клонирования в них относительно больших фрагментов ДНК, не подвергающихся внутренним перестройкам.
21. Организация фага λ. Принцип создания векторов на основе фага λ (векторы внедрения и векторы замещения).
А) Организация фага λ.
Двунитевая линейная ДНК состоит из 48514 п.н и содержит около 60 генов. На ее конах имеются однонитевые комплементарные участки, состоящие из 12 нуклеотидных остатков (cos-сайты). Они обеспечивают превращение линейной молекулы ДНК в кольцевую. Такие концы молекул получили название липких.
Ранние гены- обеспечивают копирование, транскрибирование.
Поздние гены – упаковка вирусов, синтез капсида.
Подробнее↓
Геном фага λ можно разделить на три основные части:
· Левая часть включает все гены (от Nu1 до J), белковые продукты которых необходимы для формирования капсидов и упаковки в них молекул фаговой ДНК.
· Центральная часть, расположенная между генами J и N, область генома содержащая гены, участвующие в общей рекомбинации фага (redα, и redβ), сайтспецифической интеграции ДНК фага в бактериальную хромосому (int) и исключении профага из хро-мосомы (xis). Сайтспецифическая рекомбинация идет по особым участкам на ДНК фага (att) и бактериальной хромосомы.
· Правая часть генома фага λ содержит все остальные контролирующие элементы, к которым, в частности, относятся гены, необходимые для репликации фаговой ДНК (О и Р) и для лизиса клеток (S и R).

Б) Принцип создания векторов на основе фага λ (векторы внедрения и векторы замещения).
· Вектора внедрения - несут один сайт узнавания для избранной рестриктазы, поэтому у них, как и у плазмидных векторов, длина рекДНК равна сумме длин вектора и клонируемого фрагмента.
· Векторы замещения – имеют два сайта узнавания для используемой рестриктазы, поэтому в такие векторы клонируемые фрагменты ДНК вставляют вместо участков, ограниченных данными сайтами.
· Вектора-хароны – содержат >1 различных сайтов(уникальных). Могут быть как замещения, так и внедрения.
В первом и втором случаях в реакциях образования рекДНК участвуют два фрагмента λ ДНК (левое и правое плечи векторного генома, имеют на одном из своих концов cos-сайт) и фрагмент чужДНК. Под действием лигазы благодаря cos-сайтам объединяются разные плечи λ ДНК. Затем образовавшиеся векторные молекулы реагируют с чужДНК и друг с другом, формируя конкатемеры (это субстрат при сборке фаговых частиц in vitro).
При работе с векторами замещения после их расщепления рестриктазами возникает проблема с буферными фрагментами, которые в процессе клонирования могут снова вставляться в вектор вместо чужДНК, уменьшая тем самым выход рекДНК. Проблему решают очисткой векторных плеч:
1) Используя препаративный электрофарез в 0,5%-ном агарозном геле.
2) Центрифугируя в градиенте плотности сахарозы или NaCl.
22. Понятия максимальной и минимальной емкости векторов, сконструированных на ос-нове фага λ.
Емкость фагового вектора – теоретически наибольший размер чужеродной ДНК, вставляемый в него, вычисляется как разность между максимальной величиной молекулы ДНК, упаковываемой в головку фага λ (равняется 52тпн), и минимальной величиной генома фага, необходимой для его развития. Последняя рассчитывается как разность между длиной ДНК (48,5 тпн) и суммарной длиной областей, несущественных для развития фага. К ним относится область в центре генома между генами J и N размером 16 тпн и область nin между генами P и Q размером 3,5 тпн. Таким образом, минимально возможный размер векторов (сконстр на основе лямбда) – 29 тпн (48,5 – 16 – 3,5), а максимальная величина вставки 23 тпн (52-29)
Реальная емкость вектора определяется не только его размером, но и минимальной величиной ДНК, которая может упаковываться в фаговую головку (35 тпн). Поэтому вводятся понятия максимальной и минимальной емкости фаговых векторов. Под этими понятиями подразумевается макс и мин количество ДНК, которое можно клонировать в векторе. Эти величины определяются, как разность между мин и макс размером упаковываемой ДНК и суммарным размером плеч вектора.
Напр: если у вектора сумма плеч равна 34 тпн, то в нем можно клонировать фрагменты ДНК от 2 (мин емкость) до 18 (макс)
|
из
5.00
|
Обсуждение в статье: Принцип организации фагмид (например, pBluescript). |
|
Обсуждений еще не было, будьте первым... ↓↓↓ |

Почему 1285321 студент выбрали МегаОбучалку...
Система поиска информации
Мобильная версия сайта
Удобная навигация
Нет шокирующей рекламы